 2015-05-06
2015-05-06 832
832ПРЕДИСЛОВИЕ
Б предлагаемой книге разбирается один из наиболее актуальных вопросов общей теории развития, который затрагивался мною еще четверть века тому назад («Организм как целое», 1938) и обсуждался в ряде дальнейших работ (Шмальгаузен, 1945, 1961).
Основные идеи во всех этих работах одинаковы: движущие силы индивидуального развития создаются по мере дифференци-ровки зародыша в результате взаимодействия продуктов этой дифференцировки. Взаимодействие разных частей ведет к новым дифференцировкам и дальнейшим взаимодействиям. Устойчивость организации покоится не на прочности каких-либо структур, а на сложности системы взаимозависимостей (корреляций) и на регуляторном их характере. В результате этих взаимозависимостей организм развивается на всех стадиях как одно согласованное целое. Несмотря на некоторое различие в терминологии общие установки автора не изменились.
Вместо прежнего, недостаточно определенного термина корреляция, который понимается иногда статически как сосуществование или соотношение, я употребляю теперь гораздо более выразительный русский термин «взаимодействие». Кроме того, я делаю попытку рассмотреть всю проблему органических регуляций в свете общей теории регулирующих устройств (кибернетики). Такой подход позволяет понять механизм регуляции гораздо полнее и ставит ряд новых задач перед эмбриологами. Изложение я пытался сделать возможно более доступным, но все же книга рассчитана на биологов — студентов и аспирантов, а также начинающих научных работников.
Задачей настоящей работы было не составление еще одной сводки по проблемам индивидуального развития, а освещение Уже известных фактов под несколько новым углом зрения. Поэтому в работе имеются лишь примеры, служащие для иллюстрации некоторых положений, и нет ни обстоятельных описаний, ни ссылок на литературу. Допущены также известная схематизация и упрощение сложных явлений, в которых всегда подчеркиваются лишь наиболее существенные моменты. В конце книги приложен список основной литературы, по которой читатель сможет ознакомиться более обстоятельно с разбираемыми вопросами.
230 Регуляция формообразования в индивидуальном развитии
В подготовке моей рукописи к печати мне много помогли мои сотрудники, в особенности И. М. Медведева, С. М. Эпштейн и О. П. Ольшевская, которым я здесь выражаю мою самую искреннюю благодарность.
ВВЕДЕНИЕ. ФОРМЫ РЕГУЛЯЦИИ
Одним из самых замечательных свойств живых существ является их приспособляемость к меняющейся обстановке. Приспособительные изменения захватывают как форму, так и функции живого существа. Они выражаются в изменении внешних признаков, форм поведения, соотношения частей и физиологических процессов, протекающих в органах, тканях и клетках всего организма. Живые существа находятся в процессе непрерывной перестройки и все же остаются себе подобными. Почти все изменения приспособительны, и почти все приспособительные изменения оказываются обратимыми.
Многие внешние влияния, несомненно, нарушают нормальное течение жизненных процессов. Живые существа способны, однако, в большей или меньшей мере к восстановлению своей нормальной жизнедеятельности. В большинстве случаев они справляются даже с механическими повреждениями, а иногда способны восстановить удаленные органы или даже всю свою организацию из небольшого ее участка. Все это относится, конечно, не только к взрослому организму, но в не меньшей, а обычно и в еще большей мере к развивающемуся зародышу, личинке, молодому животному или растению.
Во всех этих случаях говорят о способности живых существ к регуляции своей морфологической структуры и соотношений с внешней средой. Несмотря на разнообразные выражения явлений регуляции все они имеют общую основу, сходное происхождение и некоторые общие способы их осуществления. Способы осуществления регуляции оказываются сходными с механизмами, применяемыми в технике. Чтобы это было ясно, мы начнем с определения понятия регуляции, а затем перейдем к анализу общих основ явления и к вопросу об историческом происхождении регуляторных механизмов.
Под регулированием в широком смысле понимают действия, направленные на поддержание системы в определенном состоянии. В частности, понятие регуляции включает поддержание заданного или подходящего движения, его направления и скорости (течения, передачи энергии, химических реакций и т. п.), под-
Введение. Формы регуляции 231
деряшу заданных соотношений в этом движении и поддержание стационарного состояния и соотношений, установившихся в результате регулируемого движения (т. е. подвижного равновесия). В технике различают регулирующие механизмы следующих типов:
1) простое регулирование по заданной программе (например, направления движения корабля по ориентирам заданного курса),
2) прямое регулирование с учетом факторов, вызывающих уклонение от программы (например, управление кораблем с поправкой из расчета на его снос течением или ветром),
3) регулирование по замкнутому циклу с обратной связью, т. е. с проверкой результатов регулирования по сравнению с заданной программой и внесением соответствующих поправок (например, работа автоматического рулевого управления).
Последняя высшая форма регулирования включает и первые механизмы, т. е. программное регулирование с учетом возмущающих влияний. Если цикл регуляции с обратной связью соединен с другим подобным циклом, контролирующим и выправляющим работу первого, то такое регулирующее устройство называется самонастраивающимся.
В процессах индивидуального развития можно установить существование тех же принципов регулирования:
1) развитие по программе, заданной наследственным материалом, т. е. внутренними факторами развития,
2) развитие соответственно положению во внешней среде, т. е. в зависимости от внешних факторов,
3) регуляция в собственном смысле, т. е. выправление уклонений и восстановление «нормальных» соотношений при их нарушении. Последнее возможно лишь через посредство замкнутого цикла зависимостей, т. е. при наличии обратной связи между развивающейся частью и наследственной основой форм реагирования.
Из приведенных определений ясно, что принятое в технике понятие регулирования в применении к развивающемуся организму равнозначно всему понятию развития. Провести какую-либо грань между способами регулирования формообразования в индивидуальном развитии невозможно, так как обычная в этом случае регуляция с обратной связью, несомненно, всегда включает и оба способа выражения прямой регуляции. С другой стороны, наличие механизмов регуляции по унаследованной программе и Регуляции соответственно внешним влияниям ясно выступает у всех организмов как основа видимого процесса индивидуального развития. Регуляция с обратной связью является более совершенной надстройкой, которая усложняется и совершенствуется по мере прогрессивной эволюции организмов. Обычно такие циклы образуют в организмах все более сложные самонастраивающиеся системы регуляции. Эти последние формы регуляции соответствуют
232 Регуляция формообразования в индивидуальном развитии
обычному пониманию регуляции в живых организмах и являются основной темой дальнейшего изложения.
Регуляций физиологических функций организмов мы здесь касаться не будем. Общие принципы их осуществления те же самые и, хотя они достигают также очень большой сложности, они гораздо лучше изучены, чем формообразовательные регуляции. Последние анализируются с помощью очень трудоемких микрохирургических и других специальных методов. Биохимические их основы мало доступны для исследования вследствие ничтожных количеств действующих веществ и трудности их выделения. Однако в последнее время и здесь достигнут значительный прогресс.
Формообразовательные регуляции могут быть расчленены на частные процессы, но вообще достигают большой сложности во взаимодействующих системах со взаимным контролем результатов. Формы взаимодействия частей формообразовательных систем могут быть различными. В своем взаимодействии части системы могут оказывать друг на друга стимулирующее влияние (положительная обратная связь). Это ведет к согласованному последовательному развитию всей системы. Возможно, однако, и такое соотношение, когда одна часть оказывает на другую стимулирующее воздействие, а вторая — на первую —-задерживающее (отрицательная обратная связь). Это ведет к установлению стационарного состояния, т. е. некоторого равновесия в данной системе.
Глава I
ДЕТЕРМИНИРОВАННОЕ РАЗВИТИЕ И РЕГУЛЯЦИЯ
Рассмотрим некоторые примеры регуляции формообразовательных процессов.
1. Развитие по унаследованной программе
Такой тип регуляции легко устанавливается хотя бы при сравнении развития икры разных видов рыб в одном водоеме (или сосуде) в совершенно одинаковых условиях или при сравнении развития яиц различных птиц в одном и том же инкубаторе. В этих случаях развиваются мальки рыб и выклевываются птенцы со всеми характерными особенностями того вида, от которого получены зрелые и нормально оплодотворенные яйца. Ясно, что развитие этих особенностей связано с происхожде-
i
Глава 1. Детерминированное развитие и регуляция 233
вием яиц и определяется наследственными их свойствами. Тот j#e опыт можно произвести и с наследственными уклонениями (мутациями) и даже с уродствами, если только они •жизнеспособны. В одинаковых условиях развития их потомство вполне закономерно отличается от «нормальных» особей. «Программа» развития оказывается у мутаций определенным образом измененной, и эта измененная программа передается По наследству и дальнейшему потомству (по определенным законам).
Как развертывается развитие по унаследованной программе, можно иногда и непосредственно наблюдать уже на первых
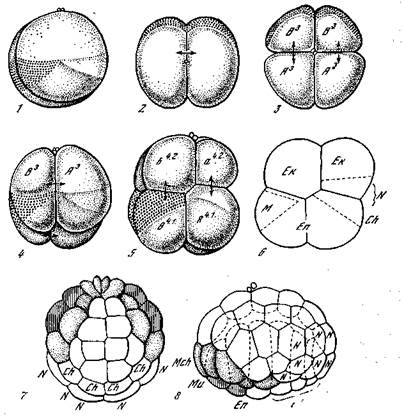
* ис. 2. Дробление асцидий. Распределение материала при дроблении яйца Ще1а [Кюн,1955]
I __ г
9 первое, второе и третье деления дробления; б — схема дробления; 7 — 64-леточная стадия; «— 76—112-клеточный эмбрион, вид с левой стороны; А, В, а, ~~ обозначения бластомеров; Ch — материал хорды; Ek — материал эктодермы;
"-материал энтодермы; М —материал мезодермы; Mch — материал мезенхимы; и —материал миобластов; JV —материал нервной системы
234 Регуляция формообразования в индивидуальном развитии
стадиях развития оплодотворенного яйца. Это касается в особенности яиц многих червей, особенно высших, моллюсков, асцидий (рис. 1) и некоторых других животных. В готовом яйце асцидии (Styela) можно различить внутреннюю массу желтка и поверхностный слой цитоплазмы, содержащей желтые пигментные зерна. Ядро яйца, располагающееся вначале в середине желтка, перемещается вместе с окружающей его прозрачной цитоплазмой к поверхности яйца, где происходят деления созревания. Одновременно с первым делением созревания прозрачная цитоплазма растекается по поверхности в направлении к экватору яйца. Непосредственно после оплодотворения возникают потоки цитоплазмы, перераспределяющие все содержимое яйца. Желток перемещается к одному полюсу (анимальному), а желтый пигмент — к другому. Яйцо сперматозоида сближается с ядром яйца, и они оба вместе с окружающей прозрачной цитоплазмой передвигаются в направлении к экватору, под которым концентрируется желтый пигмент, серпообразно охватывающий половину яйца («желтый полумесяц»). После первого деления дробления яйца происходят дальнейшие перемещения — желток перетекает обратно к нижнему, вегетативному полюсу, прозрачная цитоплазма, собравшаяся вокруг ядер, перетекает к верхнему — анимальному полюсу. Под экватором, на стороне противоположной желтому полумесяцу, концентрируется светло-серая цитоплазма в виде подобного же серого полумесяца.
После образования желтого полумесяца, яйцо, имевшее раньше радиально-симметричную организацию, получает двустороннюю симметрию. Веретено первого деления дробления становится перпендикулярно к плоскости симметрии, и соответственно первая плоскость дробления разделяет яйцо по плоскости симметрии на две совершенно равные половинки (первые два бластомера), содержащие каждая по половине от каждого полумесяца. Эти половинки яйца зеркально-симметричны и дают начало правой и левой половине тела. По мере дальнейших клеточных делений яйцо дробится последовательно на 4, 8, 16, 32, 64 бластомера и т. д. При этом происходит совершенно определенное распределение плазменных субстанций яйца по бластоме-рам, и в дальнейшем развитии эти разные бластомеры дают начало различным, вполне определенным, органам животного. Так, на стадии 32 клеток (бластомеров) в вегетативной половине яйца 6 клеток получают желтую цитоплазму яйца и дадут впоследствии начало мезодерме (т. е. мезенхиме ж мускулатуре), остальные клетки вегетативного полюса содержат желток и дадут начало энтодерме (главным образом, кишечнику). Клетки верхней»; анимальной, половины яйца дадут начало эктодерме (главным образом покровам). На стадии 64 клеток в анимальной полови0е яйца имеются 26 эктодермальных клеток, содержащих прозрачну10 цитоплазму (с очень малым количеством желтка), и 6 клеток
Глава I. Детерминированное развитие и регуляция 235
будущей нервной пластинки. В состав последней входят, кроме того, 4 клетки вегетативной половины. Все 10 клеток нервной пластинки (зачатка всей нервной системы) богаты цитоплазмой и содержат вещество серого полумесяца. Под нервной пластинкой помещаются 4 богатых желтком клетки вегетативной половины, содержащие также вещество серого Полумесяца. Это зачаток центральной скелетной оси (хорды) личинки. В вегетативной половине яйца имеются сверх того 10 богатых желтком энтодермальных клеток (зачатки кишки), 10 светло-желтых клеток будущей мезенхимы (промежуточная ткань дающая, между прочим, начало кровеносной системе) и 4 темно-желтых клетки — образователи будущей мускулатуры. Последние 14 клеток получают субстанцию желтого полумесяца, богатую митохондриями, содержащими окислительные ферменты. Во время дальнейшего развития все клетки сохраняют строго симметричное расположение и дают начало совершенно определенным частям зародыша и личинки. На рис. 1 и 2 показано распределение субстанций яйца по продуктам его дробления и дальнейшее развитие организации личинки асцидий. Это развитие связано, следовательно, с закономерным перемещением и распределением определенных веществ, содержащихся уже в плазме зрелого яйца. Правда, исследования показали, что видимые субстанции — желток, желтый и серый пигменты — являются лишь индикаторами, отмечающими известные области яйца, но вовсе не определяющими направление их развития. Дифференциальное развитие частей связано с различным состоянием и составом цитоплазмы, содержащей особенно в поверхностных, более плотных слоях некоторые более активные вещества. Эти вещества определяют ход развития соответствующих клеток, а одновременно способствуют и распределению более пассивных субстанций, используемых при нормальном развитии соответствующих органов.
Развитие, связанное с таким распределением формообразовательных веществ называют «мозаичным» (по аналогии с созданием мозаичных картин путем распределения цветных камней), или «детерминированным» (т. е. предопределенным). Оно является ярким примером простого развития по унаследованной программе. Однако и в таких случаях всегда имеются хотя бы некоторые элементы истинной регуляции, т. е. частичного восстановления нормы при ее нарушениях. При искусственном Удалении бластомеров, содержащих материал будущих органов, в развивающейся личинке эти органы полностью отсутствуют. Можно отделить друг от друга оба первых бластомера. каждый из них развивается самостоятельно в половинную личинку, содержащую одни только правые или одни только левые органы. Однако такие личинки могут принять почти нормальную внешность, так как половинки непарных органов перекраиваются: эктодерма одной стороны разрастается и покрывает
236 Регуляция формообразования в индивидуальном развитии
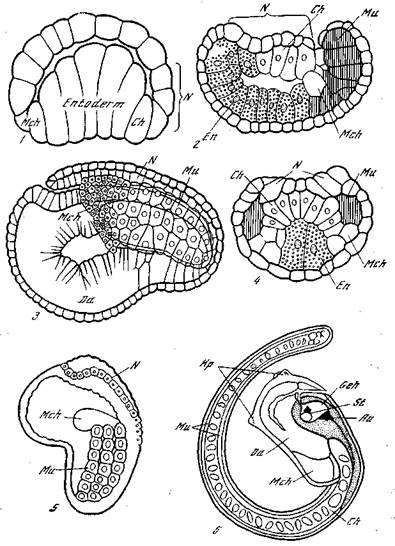
Рис. 2. Развитие асцидий [Кюн, 1955]
1 — медианный срез 76—112-клеточного эмбриона Styela partita (ср. рис. 1, 8); 2 я 4, — стадии гаструляции у Giona intestinalis (2 — срез немного латеральнее медианной плоскости; 4 — поперечный срез); 3 — нейруляция у Glavelina rissoana, начинающееся обособление туловища и хвоста; 5 — более поздняя нейрула; 6 — личинка Ascidiella scabra; Au —глаз; Ch — хорда; Da — кишка; Еп — энтодерма; Geh-~ мозг; Нр — присоски; Mch — мезенхима; Ми — мускульные, клетки; N — закладка нервной системы; St — статолит
Глава I. Детерминированное развитие и регуляция 237
дефектную сторону, половина нервной пластинки и половина энтодермального зачатка замыкаются в нервную и кишечную трубки. Следовательно, некоторая регуляция (в узком смысле) все же возможна.
При мозаичном развитии судьба каждой части определяется внутриклеточным материалом, полученным в процессе дробления из цитоплазмы яйца. Однако одновременно имеются и некоторые формообразовательные взаимовлияния. Нормально на переднем конце тела за счет его эктодермы развиваются три присоски, служащие для прикрепления личинки к субстрату перед ее преобразованием в сидячую асцидию. На изолированной эктодерме присоски никогда не развиваются. В отсутствие энтодермы, как и в отсутствие нервной трубки, присоски также не развиваются. Если повернуть анимальную половину зародыша (в которой нормально развиваются присоски) на 180° по отношению к вегетативной, то присоски развиваются совершенно нормально на переднем конце тела, а не на заднем. Это доказывает, что материал закладки не предопределен заранее в передней части эктодермы (она ведь была повернута назад, и, следовательно, присоска должна была бы развиться сзади). Оказывается присоски могут развиваться в любой части эктодермы, вступающей в определенную связь с закладками кишечника и нервной системы. В этом выражается значение взаимовлияния зачатков, которое особенно характерно для регуляционного типа развития. Это показывает, что развитие организма не бывает совершенно строго предопределенным одним только распределением субстанций, содержащихся в зрелом яйце. Известное значение всегда имеют и взаимовлияния различных частей зародыша. У зародышей асцидий и некоторых других животных эти взаимовлияния не играют большой роли и способность к регуляции у них невелика. Тем интереснее отметить, что в противоположность зародышам взрослые асцидий обладают чрезвычайно высокой способностью к регуляции в случае повреждений и даже к восстановлению всей своей организации из отдельных частей.
2. Развитие соответственно условиям внешней среды
Влияние факторов внешней среды на ход индивидуального развития сказывается в той или иной степени у всех организмов. Однако у растений это влияние выражается особенно яРко, и зависимые от этого изменения иногда резко бросаются в глаза. Так, листья многих болотных и водяных растений приобретают совершенно различный вид и строение в зависимости от водной или воздушной среды, в которых они разви-в&к>тся,
238 Регуляция формообразования в индивидуальном развитии.
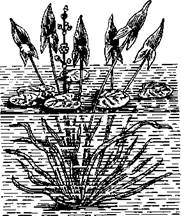
У водяного лютика (Ranunculus delphinifolius) развиваются две совершенно различные формы наземно-воздушных листьев и водных. У стрелолиста (Sagittaria) наблюдаются три формы листьев: стреловидная (развивается на суше), овальная (развивается в виде плавающих листьев на поверхности воды) и лентовидная (развивается в глубине воды) (рис. 3). Поверхность водных листьев обычно увеличенная, и
|
| Рис. 3. Изменение формы листьев на одном и том же экземпляре стрелолиста Sagittaria sagittifolia в зависимости от условий среды [Шмалъгау~ ген, 1946] |
этим они напоминают «теневые» листья наземных растений, отличающиеся большей поверхностью и нередко большим расчленением от «световых» листьев. И действительно, оказывается, водные листья развиваются именно под влиянием ослабленного освещения в воде, а не в прямой зависимости от водной среды. Вполне понятно, что как раз свет играет особенно важную роль в формообразовании растений. При этом влияет не только интенсивность света, но и длительность светового дня. Растения длинного дня недоразвиваются в условиях короткого дня и наоборот. У одного и того же растения могут развиваться в условиях длинного дня одни листья и в условиях короткого дня — другие. От этих условий зависят и форма вегетации, и цветение растений.
Все эти реакции кажутся удивительно целесообразными. Они отвечают климатическим условиям тех местностей, в которых данное растение нормально произрастает. Они как будто полностью определяются внешними факторами. Это впечатление, однако, обманчиво. Во всех случаях каждая реакция оказывается типичной для данного вида растений, и нередко даже очень близкий другой вид дает в тех же условиях среды совершенно другую реакцию. Водные листья водяного лютика характерны именно только для этого вида. Другие виды лютиков не образуют таких листьев. Теневые листья круглолистного колокольчика (Campanula rotundifolia), образующие нормально прикорневую розетку, характерны именно для этого вида, а не для других колокольчиков.
У животных также известны формообразовательные реакций на изменения во внешних факторах. У соленоводного рачка (Artemia salina) в зависимости от концентрации соли изменяет-
Глава 1. Детерминированное развитие и регуляция 239
сЯ форма и вооружение последних члеников брюшка. Однако даже у разных рас этого рачка степень реагирования на опреснение или насыщение воды солью весьма различна: каждая раса имеет свой оптимум солености воды, в котором брюшко достигает полного развития. Различно реагируют на внешние факторы и разные мутации одного и того же вида растений или животных. Это касается и таких признаков, как яровость, озимость растений или оптимальная для них длина светового дня. У животных реакции разных мутантов хорошо изучены на стадиях личиночного развития у плодовой мушки дрозофилы. Опыты показывают удивительное многообразие форм их реагирования на температуру, влажность среды и другие внешние факторы.
Во всех этих случаях, хотя факторы внешней среды и вызывают изменения в развитии, характерные особенности этих изменений, их специфика определяются наследственными свойствами данного организма (и даже отдельной особи) и не могут быть вызваны у другого вида организмов, обладающего другой нормой реакций.
И в этих случаях мы имеем, следовательно, развитие по унаследованной программе, в которой уже заложена и способность к различным формам реагирования на внешние факторы. Они образуют каждый раз характерные для данного вида организмов (точнее для данной наследственной основы — генотипа), ему одному свойственные модификации его внешнего вида и строения (т. е. его фенотипа).








