 2015-05-06
2015-05-06 376
376Типичная последовательность расчленения зачатка нервной системы определяется, по-видимому, также некоторым взаимодействием. Для нормальной организации зачатка нервной системы требуется, прежде всего, известная длительность взаимодействия эктодермы с хордомезодермой. В эктодерме сначала индуцируется лишь способность к развитию нервных тканей, а затем постепенно определяется способность к образованию нервной трубки и, наконец, детерминируются определенные отделы головного и спинного мозга. Иногда различают две формы индукции — эвокацию, т. е. влияние, вызывающее образование зачатка, и индивидуацию, т. е. влияние, вызывающее региональные различия и общую организацию зачатка. На конкретном материале трудно разграничить эти процессы, и вряд ли между ними имеются принципиальные различия. Весь процесс детерминации идет под влиянием контакта с тем же индуктором.
На стадии гаструлы при повороте эктодермы будущей нервной пластинки на 180° она образует все же под влиянием индуктора нормальную последовательность отделов мозга. На стадиях ней-руляции (закладка нервной пластинки) при таком же опыте образуется обратная последовательность отделов. Последовательность расчленения устанавливается, следовательно, одновременно с окончательной детерминацией материала нервной пластинки, очевидно, под влиянием того же индуктора. В таком случае нужно ожидать, что индуцирующие свойства различных отделов хор-Домезодермы различны. Это предположение показано экспериментально как в опытах с гомопластическими пересадками (рис. 12), так и в опытах с гетеропластической эксплантацией разных отделов хордомезодермы (рис. 13). В последнем случае для эксперимента были взяты части хордомезодермы одного вида тритонов и
256 Регуляция формообразования в индивидуальном развитии
большие куски индифферентной еще эктодермы ранней гаструлы другого вида, отличающегося по размерам и пигментации клеток. Куски хордомезодермы завертывались в эктодерму, и такая искусственная система культивировалась в солевом растворе. В результате развивались более или менее сложные образования с самыми различными органами, происхождение которых можно было точно проверить по характеру клеток. При эксплантации в такой системе материала верхней губы бластопора или образовавшейся из нее передней части хордомезодермы этот материал дифференцируется в переднюю (головную) кишку, хорду, мускулатуру. В прилежащем эктодермальном покрове (другого вида)
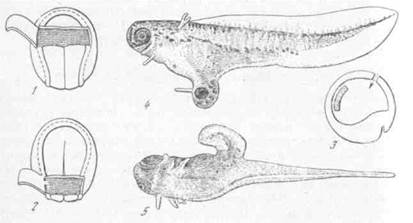
Рис. 12. Действие головного и хвостового индукторов [Кюн, 1955]
1 — 3 — схемы операций; 4 — действие головного индуктора, операция по схеме 1-+3; 5 — действие хвостового индуктора, операция по схеме 2-*-3
развиваются части головного мозга, глаза, обонятельные ямки и иногда слуховые пузырьки, т. е. только типичные головные органы. При эксплантации кусочка задней части хордомезодермы из нее в таком же опыте развиваются хорда и мускулатура, а в прилежащей эктодерме только зачаток спинного мозга, т. е. туловищные органы. В связи с концом задней части развиваются и более или менее хорошо выраженные хвосты с продолжением хорды и мускулатуры. Следовательно, передняя и задняя части индуктора действительно различаются по своим свойствам. Дальнейшие опыты показали, что между разными отделами хордомезодермы нет качественных различий. Имеются лишь какие-то количественные различия и постепенное изменение свойств при переходе от переднего конца крыши первичной кишки к заднему. Быть может, эти различия связаны с последовательностью в развитии самого материала. Передний конец образуется из того
Глава II. Регуляция формообразования у земноводных
материала верхней губы бластопора, который ранее всего ввертывается внутрь и проделывает довольно длинный путь под эктодермой до района анимального полюса. Задний конец крыши образуется позже всего — в самом конце гаструляции.
Таким образом, мы приходим к вероятному выводу о том, что региональное расчленение зачатка нервной системы зависит, по крайней мере отчасти, от свойств самого индуктора, который также обнаруживает региональные различия. Однако в индукторе эти различия имеют количественный характер постепенного изменения от переднего конца к заднему. По-видимому, такой характер различий определяется изменением концентрации каких-то
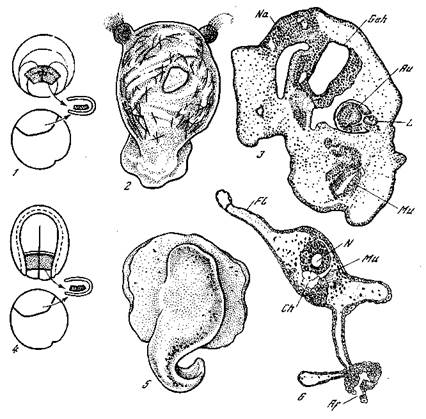
Р^с. 13. Эксперименты с помещением испытуемого индукционного материа- л«в презумптивную эктодерму [Кюн, 1955]
2> 4 — схемы операций; 2, 3 — результаты операций по схеме 1; 2 —головной «организатор» тритона в эктодерме жерлянки; 3 — головной «организатор» жерлянки в эктодерме жерлянки; 5, 6 —результаты операции по схеме 4\ 5 — туловищный Организатор» жерлянки в эктодерме тритона; 6 — то же, в разрезе; Af — задняя кищКа; Аи — глазной бокал; Ch — хорда; Л—плавник; Geh— мозг; L — хруста-Лик; Ми — мускулатура; IV — нервная трубка; Na — эпителий носовой ямки
258 Регуляция формообразования в индивидуальном развитии
веществ или изменением процессов клеточного обмена. Между тем, в зачатке нервной системы под влиянием этих количественных различий наступает разделение на качественно различные отделы центральной нервной системы (развитие которых определяется пороговым уровнем поступления индуцирующих веществ). Формообразовательное значение количественных различий в распределении известных веществ, связанное с изменением метаболизма в известных направлениях, установлено уже во многих случаях. Постепенное падение или нарастание концентрации некоторых веществ, проявлений метаболизма, индуцирующих свойств или способности к известным реакциям (компетенции) характерно для очень многих зачатков и обозначается как падение или нарастание некоторого градиента. Известный американский исследователь Чайльд пытался построить на явлениях градиента законченную общую теорию развития. Это вряд ли возможно. Однако огромное значение градиентов совершенно бесспорно. Между прочим, именно наличие градиента дает прекрасное объяснение высокой способности к регуляции всех ранних, но уже детерминированных в общем зачатков органов, так как при удалении части зачатка общий градиент не нарушается и вновь определяет развитие целого.








