 2015-05-06
2015-05-06 1074
1074Прежде всего была доказана весьма активная роль клеток верхней губы бластопора (т. е. области серого полумесяца яйца). Регуляционная способность зародышей тритонов очень высока, и у них можно получить две совершенно нормальные личинки (и взрослых животных) не только путем обособления первых бластомеров яйца (рис. 8), но и путем разделения молодого зародыша (бластулы или молодой гаструлы) на две равные части. Это достигается перешнуровкой яйца петлей из тонкого волоса. Успех операции обеспечивается, однако, лишь в том случае, если волос пересек верхнюю губу бластопора, так что каждая половина зародыша получает часть этой губы (рис. 9). Уже это показывает значение верхней губы бластопора для формообразования. Она получила даже неудачное название «организатора», так как именно от этого клеточного материала начинайся как будто процесс организации всего зародыша. Многократ-йо производились опыты пересадки кусочка верхней губы бластопора на другое место, например, на бок другой бластулы или гаструлы (рис. 10). Результат получался совершенно однозначный. Всегда клетки пересаженного кусочка (конечно,
248 Регуляция формообразования в индивидуальном развитии
в случав приживления) активно врастают внутрь и увлекают с собой также прилежащие клетки эктодермы. Они образуют непрерывный слой клеток, перемещающийся под эктодермой вперед по направлению к анимальному полюсу. В этом слое клеток посредине развивается добавочная хорда, по бокам добавочные сегменты осевой мускулатуры, почечные канальцы, а затем и другие органы. Под ними развивается иногда добавочная кишечная полость. В эктодерме на месте ее контакта с ввернувшимся
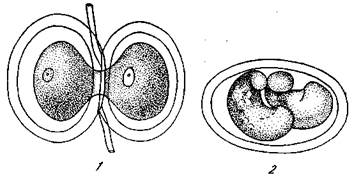
Рис. 8. Перешнуровка дробящегося яйца тритона [Дюркен, 1929]
1 — разделение бластомеров на стадии незавершенного первого деления (стадия «гантели»); 2 — получившиеся два эмбриона-близнеца в общей яйцевой оболочке
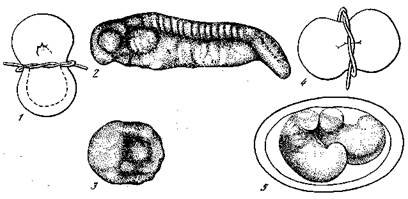
Рис. 9. Перешнуровка оплодотворенного не дробящегося яйца тритона [Гексли и де Вер, 1936]
1 — яйцо тритона, разделенное в горизонтальной плоскости на дорсальную и вентральную половины; 2 — нормальный эмбрион, развившийся из дорсальной половины; 3 — бластулоподобный шар, не способный к дальнейшему развитию, образовавшийся из вентральной половины, не содержавшей области «организатора»; 4 — яйцо тритона, разделенное в сагиттальной плоскости; 5 — нормальные эмбрионы, развившиеся в результате разделения 4, при котором в каждую половину яйца попала часть области «организатора»
Глава II. Регуляция формообразования у земноводных 249
слоем клеток развивается добавочная нервная пластинка, затем завертывающаяся в трубку; сзади развивается хвост. Таким образом, на боку зародыша — хозяина — развивается добавочный, как бы паразитирующий на нем второй зародыш. Как показыва-jox опыты пересадки губы бластопора с одного вида тритона на другой, отличающийся по своим клеткам, почти вся эта организация развивается на месте из материала хозяина, а пересаженный материал образует лишь небольшую часть осевых органов
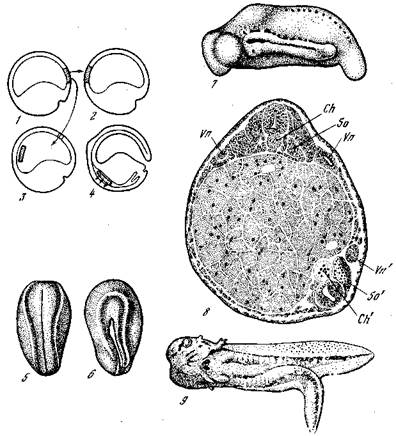
Рис. 10. Индукция дорсальной губой бластопора [Кюн, 1955]
1~~4 — схема операций; 5 — 9 — зародыши обыкновенного тритона с трансплантатами дорсальной губы бластопора; 5, 6 — нейрула хозяина (5 — вид с дорсальной стороны, нервная пластинка хозяина; б — вид с вентральной стороны, индуциро-ванная нервная пластинка); 7 — зародыш с индуцированным дополнительным за-Родышем на боку; 8 — поперечный срез того же зародыша; 9 — другой зародыш с хорошо развитым дополнительным зародышем; So' — сомит индуцированного за-Родыша; Vn — почечные канальцы хозяина; Vn'— почечные канальцы индуцирован-н°го зародыша
250 Регуляция формообразования в индивидуальном развитии
(хорды и иногда сомитов). В этом видна как будто организующая роль губы бластопора. В действительности можно отметить лишь две особенности, характеризующие клеточный материал верхней губы бластопора. Во-первых, он энергично врастает внутрь под эктодерму и дает затем начало клеткам хорды и мезодермы, как и должно бы быть по его происхождению. В этом видна некоторая способность к самодифференцированию. Во-вторых, он вовлекает в свои движения соседний эктодермальный материал, который затем приобретает все свойства типичной хордомезодермы. В этом выражается его способность к индукции. Такое явление уподобления клеток называется ассимиляторной индукцией.
Свойства материала верхней губы бластопора этим не ограничиваются. Образовав зачаток хордомезодермы и вступив в контакт с эктодермой, тот же клеточный материал вызывает в ней образование нервной пластинки. Таким образом, материал хордомезодермы не только может побудить развитие чуждого материала в том же самом направлении, в котором он развивается сам, но может вызвать в соседней части развитие совсем другого органа. Это явление также называется индукцией. В отличие от ассимиляторной индукции иногда говорят в таких случаях о гетерогенной индукции. Существование обеих форм индукции доказано очень многочисленными экспериментами. Наличие ассимиляторной индукции было подтверждено следующим красивым экспериментом: производилась пересадка меченого с помощью прижизненной окраски чуждого материала — эктодермы, взятой с боковой стороны зародыша, в область верхней губы бластопора. После того как этот материал ввернулся внутрь и вошел в состав крыши первичной кишки, окрашенный кусочек извлекли оттуда и пересадили под эктодерму другому зародышу. Здесь на новом месте контакта развилась добавочная нервная пластинка. Следовательно, чуждый материал, внесен-вый в область крыши первичной кишки, сам приобрел все свойства последней и, в частности, способность индуцировать нервную систему.
При ассимиляторной индукции материал разного происхождения вовлекается в одни и те же условия клеточного метаболизма, и пет оснований думать, что при этом происходит непременно передача каких-то специфических веществ. При гетерогенной индукции дело обстоит иначе. Трудно сказать, каково значение ассимиляторной индукции в нормальном развитии. Однако гетерогенная индукция является, несомненно, одним из основных выражений формообразовательного значения взаимодействия частей в индивидуальном развитии.
Явление индукции нервной пластинки послужило объектом исследования множества экспериментов, выполненных разными учеными. Невозможно дать полный обзор их результатов. Мы
Глава П. Регуляция формообразования у земноводных 251
остановимся только на самом существенном, относящемся к на-pieH основной теме. В процессе индукции происходит действительно стойкое преобразование материала, а вовсе не простая активация каких-либо местных свойств материала. Эктодерма сама по себе, взятая даже из спинной области бластулы или молодой гаструлы, не может дать начало чему-либо организованному, даже эпидермису. Это показывают опыты искусственного из-
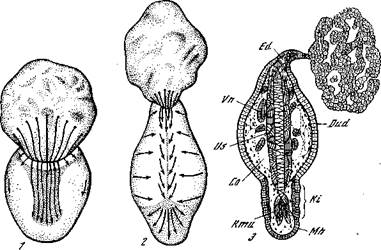
Рис. 11. Экзогаструляция у аксолотля [Кюн, 1955]
1, 2 — схемы передвижения материала; 3 — более старший экзогаструлировавший зародыш; Со — целом; Dud — тонкая кишка; Ed — хвостовая кишка; Кг — головная кишка; Кти — мускулатура головы; Mh — ротовая полость; Us — сомиты; Vn — почка
вращения процесса гаструляции (рис. 11) при помещении бластулы в гипертонический солевой раствор. Гаструляционные движения происходят в другом направлении, чем в норме. Материал верхней губы и всей краевой зоны не вворачивается внутрь, а обрастает вегетативные клетки снаружи, и это приводит к тому, что хорда образуется на поверхности, а клетки мезодермы врастают между нею и массой вегетативных клеток. После этого весь зародыш покрывается энтодермой, в которой происходят характерные дифференцировки различных отделов кишечника с железами, жаберными мешками и даже вкусовыми Точками на переднем конце. Весь этот кишечник оказывается вывернутым наизнанку (внутренней поверхностью наружу). ** мезодерме также путем самодифференцирования образуются Мышцы, полость тела и почечные канальцы. Однако в собственно ЗДтодерме, которая остается снаружи в виде обособленного
252 Регуляция формообразования в индивидуальном развитии
мешка, не происходит никакой дифференцировки. Зародыш ц0, лучается без зачатка нервной системы, так как эктодерма не могла вступить в контакт с хордомезодермой.
То же самое показывают и опыты культивирования различных частей раннего зародыша в стерильных изотонических солевых растворах (раствор Гольтфретера). Части энтодермы дают путем самодифференцирования начало тем или иным отделам кишечника соответственно их происхождению (мозаичное развитие). Однако части эктодермы тритонов совершенно не способны к какой-либо дифференцировке (в чистой культуре, т. е. без других тканей). Брюшные части краевой зоны дают в культурах путем самодифференцирования почечные канальцы, кровь и части кровеносной системы, которые нормально развиваются из латеральных частей мезодермы. Спинной отдел краевой зоны, т. е. материал, который нормально вворачивается через верхнюю губу, дает начало в средней части хорде и по бокам — осевой мускулатуре (сомитам), в изолированном состоянии хорошо дифференцируется, но не самоорганизуется. В культурах куски, даже из средней части, которые нормально дают хорду, развиваются не только в ткань хорды, но и в мышцы. Кроме того, они. дают нередко и совершенно чуждые образования — эпидермис, нервную ткань и даже части мозга. Из боковых частей того же отдела, нормально дающих начало мускулатуре тела, развиваются в изолированных культурах также куски хорды и нервная ткань, т. е., безусловно, чуждые дифференцировки. Это показывает, что в организме имеются не только факторы, стимулирующие образование разнородных дифференцировок, но и тормозящие влияния, не допускающие некоторые из возможных дифференцировок.
Ни эктодерма, ни материал верхней губы бластопора сами по себе не способны дать начало чему-либо организованному. Только во взаимном контакте решается вопрос о их закономерном расчленении и развитии. Те самые куски материала краевой зоны, которые, будучи изолированы, дают (в эксплантате) начало почти всему, чему угодно, развиваются при пересадке под эктодерму (в имплантате) исключительно в хорду и мускулатуру. То же самое происходит и в культурах (в эксплантатах), если кусочек краевой зоны культивировать в обвертке из индифферентной эктодермы. В этом случае из материалов краевой зоны развиваются хорда и мускульные сегменты тела, а в прилежащей эктодерме индуцируются и развиваются различные отделы центральной нервной системы и связанные с ними органы чувств.
Все эти опыты, так же как и многие опыты взаимных пересадок, ясно показывают значение взаимодействия между материалом хордомезодермы и эктодермы. Материал хордомезодермы (т. е. ввернувшийся материал дорсальной области краевой зоны) возбуждает клеточные перемещения в нижнем слое эктодермы,
Глава П. Регуляция формообразования у земноводных 253
иХ концентрацию и преобразование в закладку нервной системы. С ДРУГ0И стороны, нервная пластинка оказывает обратное воздействие на крышу первичной кишки, организуя ее и подавляя в ней способность к образованию чуждых для данного места образований — эпидермиса, нервной ткани. В крыше первичной киптки образуются по бокам мускульные сегменты, которые в свою очередь влияют на материал нервной пластинки и определяют по ее краям такое же сегментальное расположение нервных узлов.
Возможность тормозящего влияния можно иллюстрировать еще следующим примером. Эктодерма из бластулы лягушки образует в эксплантате неорганизованную массу. Однако в отличие от эктодермы хвостатых земноводных, в ней дифференцируется железистое образование — присоска. Последняя развивается нормально в виде парного образования на переднем конце головы. В эксплантате в каждом куске эктодермы, взятом с любого места зародыша, всегда развивается по одной присоске, иногда огромной величины. Таким образом, вся эктодерма способна дать начало присоске. Однако нормально в зародыше развивается только одна пара присосок на брюшной стороне передней части головы, под влиянием контакта со стенкой передней кишки. Последняя изолирует эктодерму от влияния мезодермы и тем самым определяет место закладки и ее парность. При отсутствии этого контакта, т. е. в остальной части тела, способность к образованию присосок подавляется подлежащей мезодермой.
2. Индукция и реакция. Детерминация и компетенция
Основной индуктор (верхняя губа бластопора и развивающаяся из этой области хордомезодерма) был назван «организатором». Это наименование преувеличивает его роль. В сущности он только определяет энергичное перемещение клеток, характер движений материала и при установлении контакта с эктодермой выбор одного из двух возможных направлений развития эктодермы: в сторону развития эпидермиса или в сторону развития нервной пластинки. Одновременно материал «организатора» сам «организуется» под влиянием развивающейся нервной пластинки. Правильнее говорить просто об индукции, причем следует помнить, что в явлениях индукции выражается результат взаимодействия известных частей, т. е. что влияние имеет двусторонний характер. Индукция проявляется во взаимодействующей системе, состоящей по меньшей мере из двух частей. Более активную часть совершенно условно называют индуктором (или эвокато-Р°м), а менее активную часть, отвечающую явной реакцией, можно назвать так же условно реактором.
Реагирующая ткань далеко не индифферентна — она способна лишь к немногим реакциям и к тому же меняется по мере
254 Регуляция формообразования в индивидуальном развитии
развития. Она способна реагировать лишь в определенное время, когда она достигает известной степени зрелости. Кроме того, она и сама оказывает обратное воздействие на индуктор или сама приобретает свойства индуктора. Последнее также доказано многочисленными опытами пересадки частей нервной пластинки.
При пересадке части верхней губы бластопора на другое место или пересадке куска хордомезодермы под эктодерму другого зародыша (на стадии бластулы или гаструлы) индукция вовсе не наступает в любое время — добавочная нервная пластинка на ненормальном месте закладывается точно в то же самое время, когда закладывается и нормальная нервная пластинка. Только в нормальное время она «компетентна» к ответной реакции. Во время уже наступившей ыейруляции, когда выявляется закладка нормальной нервной пластинки, вся остальная эктодерма теряет свою нейральную компетенцию. Возможно, что это связано с израсходованием материала нижнего слоя эктодермы для образования нормальной нервной пластинки. Пересадки губы бластопора или хордомезодермы тогда уже не могут вызвать реакции образования добавочного зачатка нервной системы.
В результате индукции наступает прочное, необратимое определение судьбы обеих частей эктодермы. После закладки материал нервной пластинки окончательно детерминирован и при любых пересадках (а также при культивировании в эксплантате) дает начало только частям нервной системы. Вся остальная эктодерма также окончательно детерминирована и при любых пересадках дает начало только эпидермису. Детерминация зачатка есть процесс, зависимый от другого зачатка (индуктора) или от наличия известных веществ внутри клеток зачатка. Детерминированные зачатки в дальнейшем развиваются уже путем самодифференцирования. Конечно, эти процессы не наступают мгновенно, они развиваются постепенно, по этапам, в течение которых детерминация меняется от лабильной, еще обратимой при изменении условий развития, до прочной, необратимой. По мере прохождения последовательных этапов детерминации меняется иногда не только количественное ее выражение, но и качественное. На стадии ранней гаструлы эктодерма спинной ее части еще недетерминирована. На стадии средней гаструлы детерминация имеет лишь весьма общий характер и ведет к развитию (в эксплантате) лишь небольших нервных образований. Позднее детерминация оказывается более полной и ведет к развитию более крупных и лучше организованных зачатков головного и спинного мозга. К концу гаструляции и в начале нейруляции части нервной пластинки уже прочно и окончательно детерминированы: они дают начало различным отделам мозга точно согласно месту их происхождения. То же самое показывают и опыты пересадки. На стадиях гаструляции отдельные куски эктодер-мы, соответствующие будущей нервной пластинке, развиваются
Глава II. Регуляция формообразования у земноводных 255
на новом месте в обычный эпидермис, позднее из них развиваются целостные закладки нервной системы, а на стадиях нейруля-дяи такие же куски нервной пластинки дают только части мозга, строго соответствующие месту происхождения трансплантата. Вначале наступает детерминация органа вообще, а позднее детерминируются его отделы.
На первых стадиях нейруляции еще можно удалить часть закладки, и из оставшейся части разовьется вполне нормальная нервная система со всеми ее отделами. На более поздних стадиях развития нервной пластинки получившиеся дефекты уже не восполняются. Таким образом, ранние закладки нервной системы (это касается и закладок других органов) способны к регуляции, и при удалении ее части развиваются все же как одно целое, в котором наступает типичное расчленение. По мере дальнейшего развития это расчленение необратимо фиксируется.

