2015-04-30
2015-04-30 10887
10887| Тема: | Фосфолипиды. Физико-химические свойства фосфолипидов. Изучение химического состава фосфолипидов |
| Цель работы: | Освоение методов идентификации и установления химического состава фосфолипидов |
Оборудование и материалы:
· Водяная баня
· Плитка электрическая
· Горелка спиртовая
· Микроскоп
· Пипетки стеклянные на 1 и 5 мл
· Микропипетки автоматические
· Цилиндры мерные на 250 и 100 мл
· Стаканы на 50 и 100 мл
· Воронки
· Пробирки стеклянные химические
· Штативы для пробирок
· Ступка с пестиком
· Тигель
· Предметное стекло
· Бумага фильтровальная
· Палочки стеклянные
· Бумага индикаторная лакмусовая
Реактивы:
· Яйцо
· Гидроксид натрия (NaOH), 10%-ный раствор
· Карбонат натрия (Na2CO3), крист.
· Нитрат калия (KNO3), крист.
· Молибденовый реактив: молибдат аммония ([NH4]2MoO4),
15%-ный раствор / HNO3(конц.) в соотношении 110 / 90
· Магнезиальная смесь: MgCl2 (10%-ный), NH4Cl (20%-ный), NH4OH
· Хлорид кадмия (CdCl2), насыщенный спиртовой раствор
· Борная кислота, кристаллическая (либо безводный MgSO4, либо безводный KHSO4)
· Эфир
· Спирт (этиловый), 96%-ный
· Ацетон
· Соляная кислота (HCl), 10%-ный раствор
· Азотная кислота (HNO3), 10%-ный раствор
· Вода дистиллированная
Теоретическая часть
Фосфолипиды
Существует несколько классов мембранных липидов. Они отличаются от триацилглицеролов тем, что наряду с углеводородными цепями содержат одну или несколько сильно полярных «головок». На этом основании мембранные липиды часто называют также полярными липидами. В наибольшем количестве в мембранах присутствуют полярные липиды, представленные фосфолипидами. Фосфолипиды являются производными либо трехатомного спирта глицерола, либо более сложного аминоспирта сфингозина. Те фосфолипиды, которые являются производными глицерола, получили название фосфоацилглицеролов. Эти соединения состоят из глицерола (в качестве основы всей структуры), двух остатков жирных кислот и фосфорилированного спирта. В фосфоацилглицеролах гидроксильные группы при С1 и С2 атомах в глицероле этерифицированы карбоксильными группами двух жирных кислот. Третья гидроксильная группа глицерола этерифицирована фосфорной кислотой. Образующееся при этом соединение является простейшим фосфоацилглицеролом и называется диацилглицерол-3-фосфатом или фосфатидной кислотой. Это соединение представляет собой ключевой промежуточный продукт для биосинтеза других более сложных фосфолипидов. Все основные мембранные фосфолипиды образуются путем этерификации фосфатной группы фосфатидной кислоты с гидроксильными группами спиртов, таких, как серин, этаноламин, холин, глицерол и инозитол. Таким образом, к фосфолипидам относятся фосфатидная кислота, фосфатидилсерин, фосфатидилэтаноламин, фосфатидилхолин, фосфатидилглицеролы и фосфатидилинозитол. Единственный фосфолипид мембран, не являющийся производным глицерола, – это сфингомиелин.
Фосфатидная кислота и фосфатидилглицеролы
Исходным соединением для всех фосфоацилглицеролов (т.е. фосфолипидов) служит, как уже упоминалось, фосфатидная кислота (рис. 11.1). В свободном виде это соединение встречается в крайне незначительных количествах. С другой стороны, фосфатидная кислота является важнейшим промежуточным соединением в процессах биосинтеза триацилглицеролов и множества фосфолипидов.
Крайне интересными фосфолипидами являются так называемые кардиолипины, которые представляют собой «двойные» фосфатидилглицеролы (рис. 11.2). Главная особенность кардиолипинов в том, что эти соединения локализуются преимущественно во внутренней мембране митохондрий.
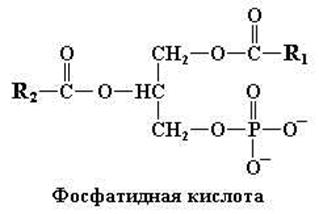
Рис. 11.1. Структурная формула фосфатидной кислоты. Второй углеродный атом (С2) глицерола в молекуле асимметричен и имеет
L-конфигурацию
Кардиолипин – фосфолипид, входящий в состав митохондриальных мембран. Он образуется из двух молекул фосфатидной кислоты, которые соединены друг с другом «мостиком», представленным остатком глицерола.
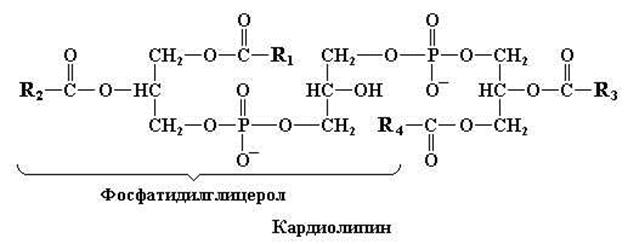
Рис. 11.2. Структурная формула кардиолипина («двойного» фосфоглицерида) содержится в больших количествах в мембранах митохондрий. Буквами R с соответствующими индексами обозначены углеводородные цепи разных длинноцепочечных жирных кислот
Фосфатидилхолин (лецитин)
Лецитины, как и нейтральные жиры, содержат глицерол и жирные кислоты, но в их состав входят также фосфорная кислота и азотистое основание – холин. Лецитины широко представлены в клетках различных тканей, они выполняют как метаболические, так и структурные функции. Большинство данных фосфолипидов, как правило, содержит насыщенный ацильный радикал в положении С1 и ненасыщенный радикал в положении С2 глицерола.
При рН 7,0 остаток фосфорной кислоты в лецитинах, а также во всех других фосфолипидах несет отрицательный заряд. Кроме того, при рН, близких к нейтральным, азотистые основания в составе фосфолипидов (рис. 11.3, 11.4, 11.6) могут нести один или несколько электрических зарядов. Таким образом, фосфолипиды содержат группировки двух разных типов, а именно полярные гидрофильные «головки» и неполярные гидрофобные хвосты. Такие соединения называют амфипатическими веществами.
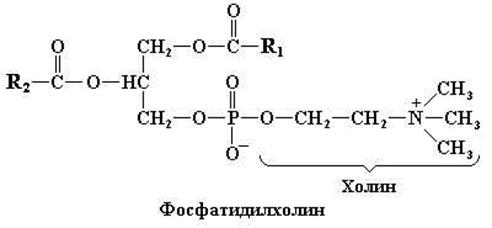
Рис. 11.3. Структурная формула наиболее известного фосфолипида – фосфатидилхолина
Фосфатидилэтаноламин (кефалин)
Кефалины отличаются от лецитинов только тем, что у них азотистое основание холин заменено другим азотистым основанием – этаноламином.
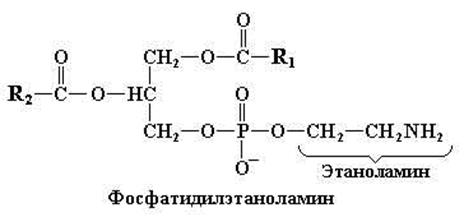
Рис. 11.4. Структурная формула фосфатидилэтаноламина
Фосфатидилинозитол
В состав фосфатидилинозитола кроме глицерола, двух остатков жирных кислот и остатка фосфорной кислоты входит шестиатомный циклический спирт инозитол (рис. 11.5). Инозитол в этом соединении представлен одним из своих стереоизомеров – миоинозитолом. Кроме собственно фосфатидилинозитола в клетках присутствует его фосфорилированная форма, представленная фосфатидилинозитол-4,5-дифосфатом, который также является важным компонентом клеточных мембран. Данное соединение обладает ярко выраженными физиологическими эффектами. В частности, фосфатидилинозитол-4,5-дифосфат принимает участие в физиологической регуляции процессов секреции, осуществляемой с помощью секретогенных соединений, которые взаимодействуют с соответствующими рецепторами на базальной стороне экзокринных клеток. Например, в результате связывания ацетилхолина или холецистокинина со своими рецепторами на базальной стороне экзокринных клеток происходит активация внутриклеточной фосфатидилинозитол-специфичной фосфолипазы С, которая расщепляет фосфатидилинозитол-4,5-дифосфат с образованием инозитол-1,4,5-трифосфата и диацилглицерола. Последние, в свою очередь, запускают высвобождение Ca2+ из эндоплазматического ретикулума в цитозол (инозитол-1,4,5-трифосфат) или активируют протеинкиназу С (диацилглицерол), что в конечном итоге обеспечивает выброс белковых продуктов из экзокринных клеток. Таким образом, оба этих соединения действуют как внутриклеточные вторичные мессенджеры передачи сигнала.
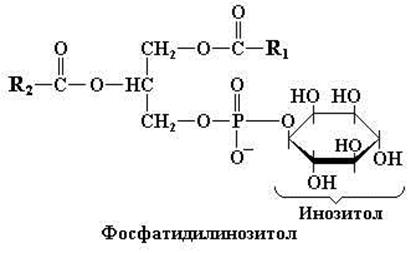
Рис. 11.5. Структурная формула фосфатидилинозитола
Фосфатидилсерин
В тканях находится также родственный кефалину фосфолипид, содержащий вместо этаноламина остаток аминокислоты серина.
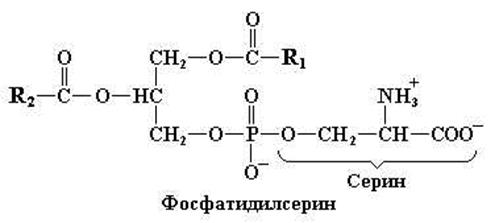
Рис. 11.6. Структурная формула фосфатидилсерина
Сфингомиелины
Сфингомиелины в больших количествах встречаются в нервной ткани. При гидролизе сфингомиелинов образуются жирная и фосфорная кислота, холин и сложный аминоспирт сфингозин (рис. 11.7).
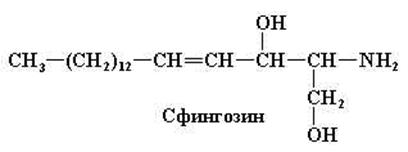
Рис. 11.7. Структурная формула сфингозина
В составе этих соединений глицерол отсутствует. При взаимодействии сфингозина с жирной кислотой (посредством амидной связи) образуется базовая структура, получившая название церамида. Церамид обнаруживается также в составе гликолипидов (рис. 11.8).
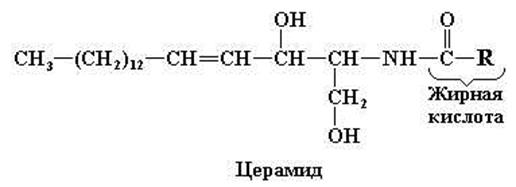
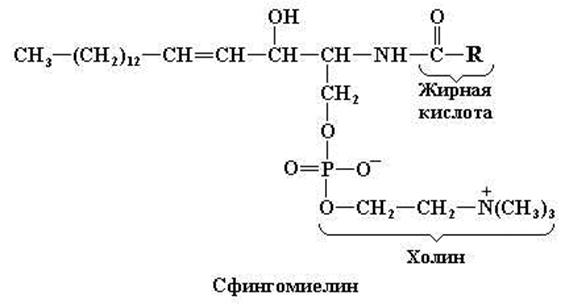
Рис. 11.8. Структурные формулы церамида и сфингомиелина
Благодаря присутствию фосфохолина в составе сфингомиелинов последние часто относят к фосфолипидам. Однако вследствие того, что гидрофобная часть этих молекул представлена церамидом, сфингомиелины более целесообразно считать сфинголипидами.
Сфинголипиды (гликосфинголипиды)
Гликолипиды широко представлены в тканях, особенно в нервной ткани, в частности в ткани мозга. Они локализованы преимущественно на наружной поверхности плазматических мембран, где их углеводные компоненты входят в число других углеводов клеточной поверхности.
Главной формой гликолипидов в животных тканях являются гликосфинголипиды. Они состоят из церамида, а также одного или нескольких остатков моносахаридов. На основании строения углеводного компонента гликолипидов их делят на цереброзиды и ганглиозиды. Двумя простейшими соединениями, относящимися к цереброзидам, являются галактоцереброзид (галактозилцерамид) и глюкоцереброзид (глюкозилцерамид). Как видно из названия этих гликолипидов, они состоят из церамида и только одного углеводного остатка. Галактозилцерамид – главный гликосфинголипид мозга и других нервных тканей, но в небольших количествах он встречается также во многих других тканях. В его состав обычно входят различные С24-жирные кислоты. Галактозилцерамид может превращаться в сульфогалактоцереброзид (сульфогалактозилцерамид) (сульфатид), который в больших количествах содержится в миелине. Простые гликосфинголипиды, входящие в состав тканей, отличных от нервной, представлены главным образом глюкозилцерамидом. В небольших количествах он имеется и в ткани мозга (рис. 11. 9).
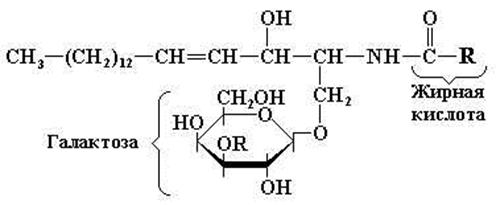
Галактозилцерамид (если R = H), cульфатид (если R = SO2-4)
Рис. 11.9. Структура галактоцереброзида и сульфогалактоцереброзида
Более сложными гликосфинголипидами являются ганглиозиды, которые образуются из глюкозилцерамида посредством присоединения к нему дополнительных остатков моносахаридов. Ганглиозиды – это гликосфинголипиды, дополнительно содержащие, кроме того, еще одну или несколько молекул сиаловой кислоты. В тканях человека доминирующей сиаловой кислотой является нейраминовая кислота, которая часто ацетилирована (см. лабораторную работу № 10). Остатки N-ацетилнейраминовой кислоты встречаются также в олигосахаридных боковых цепях некоторых мембранных гликопротеидов.
В сером веществе мозга ганглиозиды составляют около 6% мембранных липидов. Они обнаруживаются также, хотя и в меньших количествах, в мембранах клеток других (не нервных) тканей. Например, ганглиозид GМ1 является рецептором холерного токсина в эпителии кишечника человека. Ганглиозиды – важные компоненты расположенных на поверхности клеточных мембран специфических рецепторных участков. Так, они находятся в тех специфических участках нервных окончаний, где происходит связывание молекул нейромедиатора в процессе химической передачи импульса от одной нервной клетки к другой.
Наиболее простым по строению ганглиозидом, встречающимся в нервной ткани, является ганглиозид GМ3 (сокращение GМ3 означает следующее: G – ганглиозид, М – моносиаловое соединение, индекс 3 – условный номер, показывающий порядок элюции при хроматографическом разделении). Он включает церамид, одну молекулу глюкозы, одну молекулу галактозы и одну молекулу нейраминовой кислоты. Структура более сложного ганглиозида GМ1, образующегося из GМ3, показана на рис. 11.10 и на рис. 10.2 (лабораторная работа №10).

Рис. 11.10. Структура ганглиозида GМ1
Как видно, ганглиозиды GМ1 и GМ3 содержат только по одному остатку N-ацетилнейраминовой кислоты. В то же время другие ганглиозиды могут содержать от одного до пяти остатков сиаловой кислоты: их называют соответственно моно-, ди-, трисиалоганглиозиды и т.д.
Амфипатические свойства липидов
Как известно, триацилглицеролы и воска нерастворимы в воде, поскольку состоят преимущественно из гидрофобных (углеводородных) цепей. Однако жирные кислоты (в первую очередь их соли), фосфолипиды, сфинголипиды, желчные кислоты в виде солей и, в меньшей степени, холестерол содержат кроме гидрофобных «хвостов» полярные «гидрофильные» группы. Таким образом, одна часть их молекул гидрофобна (нерастворима в воде), а другая – гидрофильна (растворима в воде). Такие молекулы называют амфипатическими соединениями (рис. 11.11).
1 2
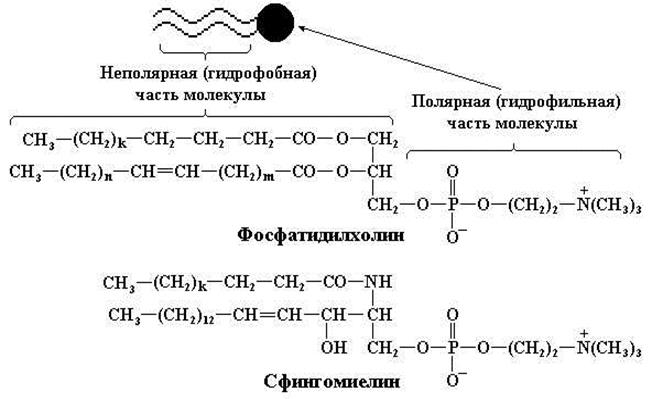
Рис. 11.11. Структура амфипатических (амфифильных) соединений фосфатидил–холина и сфингомиелина. В верхней части рисунка приводится схематическое изображение амфипатического липида:
1 – два гидрофобных «хвоста», 2 – гидрофильная «головка»
Посмотрим теперь, как ведут себя фосфолипиды и гликолипиды в водной среде. Когда амфипатические вещества смешиваются с водой, их молекулы спонтанно организуются таким способом, который удовлетворяет одновременно двум противоположным требованиям. Они организуются так, что их гидрофильные «головки» погружаются в воду в то время как гидрофобные «хвосты» в контакт с водой не вступают, а контактируют только между собой, с воздухом или гидрофобными средами, например маслом. По существу, таким путем могут образовываться три разные структуры: однослойные (монослои), мицеллы и двуслойные (бислои).
При встряхивании в воде или водных растворах фосфолипиды и гликолипиды спонтанно формируют мицеллы, в которых углеводородные «хвосты» липидов спрятаны внутри структуры, а электрически заряженные гидрофильные «головки» располагаются на поверхности частиц, взаимодействуя с водным окружением (рис. 11.12). Такие мицеллы могут состоять из многих тысяч липидных молекул. Фосфолипиды и гликолипиды способны спонтанно образовывать мицеллы.