 2015-05-13
2015-05-13 1807
1807Учебное пособие для студентов
I курса медицинских вузов
Казань, 2010
ББК 52.5
УДК 575
Печатается по решению Центрального координационно-методического совета Казанского государственного медицинского университета
Составители:
профессор кафедры медицинской биологии и генетики
Семенов Валерий Васильевич,
ассистент кафедры медицинской биологии и генетики
Харитонов Вадим Сергеевич
Рецензенты:
заведующий кафедрой генетики Казанского государственного
университета, профессор Барабанщиков Б.И.,
заведующий кафедрой генетики Казанского государственного
педагогического университета, профессор Закиев Р.К.
Хранение и реализация наследственной информации: Учебное пособие для студентов I курса медицинских вузов/ В.В. Семенов, В.С. Харитонов. – Казань: КГМУ, 2010. – 47 с.
Предназначено для студентов I курса медицинских вузов для самостоятельного изучения и получения знаний в области генетики клетки, механизмов функционирования генов, реализации заложенной в них информации. Пособие освещает вопросы хранения генетической информации и её защиты от мутагенных факторов.
© Казанский государственный медицинский университет, 2010 Содержание
Репликация ДНК …………………………………………………...…………..4
1. Определение и общие представления о репликации ДНК…….………….4
2. Периоды репликации….……………………….……………………………5
3. Коррекция (контроль) репликации ……………………………...……….12
4. Конвариантная редупликация как основа мутационной
изменчивости……………………………………………………………...….14
5. Метилирование ДНК…………………………………………..…………..14
6. Лекарственные препараты и репликация……..………………….……….15
Репарация ДНК ……….…………………………………………..………......16
1.Общее представление о репарации…….……………………….…………16
2. Виды репарации……………………………………………………………17
Общее представление о транскрипции ………………………………….…..20
1. Периоды транскрипции……………………………………………………21
2.Ошибки спаривания при транскрипции. Лекарственные препараты
и транскрипция…………………………………….….………………………25
3. Посттранскрипционные процессы. Процессинг (созревание РНК)….....27
3.1. Взаимодействие про и-РНК с белком..…………………………….28
3.2. Метилирование про и-РНК…………………………………………28
3.3. Кэпирование 5’ – конца…..…………………………..………..……29
3.4. Полиаденилирование………………………….…………….………29
3.5. Сплайсинг……………..……………………………..………...…….31
3.6. Альтернативный сплайсинг….………………………………...…...32
Трансляция ………………………………………………………….…….…...34
1. Общее представление о трансляции. Материальное обеспечение
процесса…………………………………………………..………….….…….34
2. Периоды трансляции…………..………………………….……………….37
3. Посттрансляционные процессы. Фолдинг и модификация белка……....41
Экспрессия генов. Особенности экспрессии генов у про- и эукариот …....42
1. Этапы экспрессии генов у эукариот…………………………………...…42
2. Особенности экспрессии генов у прокариот….………………………….43
3. Регуляция экспрессии гена….……………………………………….……42
3.1. Регуляция экспрессии генов у прокариот на уровне инициации
транскрипции…………………………………….……………………....43
3.2. Регуляция экспрессии генов у эукариот…….……………………..44
РЕПЛИКАЦИЯ ДНК
1. Определение и общие представления о репликации ДНК
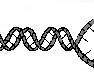
Репликация ДНК – процесс передачи информации от ДНК на ДНК в результате самовоспроизведения обеих матричных нитей всей молекулы ДНК.
Репликация характерна для ДНК и РНК. В настоящее время термин репликация чаще всего используется для обозначения синтеза новых цепей ДНК на матричных нитях. В настоящем разделе мы будем касаться только репликации ДНК.
Из школьного курса известно, что репликация у животных осуществляется в ядре клетки и митохондриях. Вначале молекула ДНК деспирализуется, нити отходят друг от друга и на каждой свободной нити синтезируется новая копия. Синтез ДНК на нитях происходит в противоположных направлениях (рис. 1). После завершения синтеза формируются две молекулы ДНК. В каждой молекуле ДНК содержится одна материнская и одна дочерняя нить, т.е. законсервирована

 3’ 5’
3’ 5’
 5’ 3’
5’ 3’
3’  5’
5’
 5’
5’  3’
3’
 3’ 5’
3’ 5’
 5’ 3’
5’ 3’
Рис. 1. Схема репликации ДНК. В каждой вновь синтезированной молекуле ДНК содержится одна нить (сплошная линия) матричной молекулы ДНК, а другая нить – вновь синтезированная
половина материала материнской молекулы ДНК. Поэтому такой синтез называют полуконсервативным. После окончания синтеза две молекулы ДНК отходят друг от друга, но остаются соединёнными в области центромер. Полностью начинают отходить молекулы ДНК друг от друга в начале профазы, когда соединяющая их центромера делится. Всё вышеперечисленное входит в понятие репликативный синтез ДНК. Этот синтез осуществляется в S-период клеточного цикла. К концу этого периода синтез прекращается. Всё вышесказанное Вы должны знать, проходя обучение по школьной программе. В предлагаемом Вам пособии мы углубим Ваше представление о репликации, показав не только молекулярное хозяйство этого процесса, но и его медицинскую значимость.
Прежде всего отметим, что учёные различают ещё один вид синтеза ДНК – репаративный. Он связан с синтезом ДНК в месте её повреждения. Повреждения уникальной структуры ДНК способны возникнуть в любой стадии клеточного цикла и в виду своей опасности должны быть немедленно восстановлены. В противном случае клетку ожидают целый ряд неприятных последствий, которые в конечном этапе приводят её к гибели. Например, если возникают обширные пов-
реждения ДНК, то они, как правило, не способны восстанавливаться и клетка гибнет. Если повреждение ДНК небольшое (а они возникают чаще всего), то оно восстанавливается достаточно быстро, синтез ДНК в месте повреждения непродолжителен и не требует больших энергетических затрат. Такой синтез носит название внеплановый синтез ДНК или репаративный синтез ДНК. Напротив, при репликации синтезируется заново вся молекула ДНК хромосомы, продолжительность её измеряется часами, при этом расходуется значительное количество энергии и заготовленного материала.
2. Периоды репликации
Для репликации (как и для всякого матричного процесса) характерны три периода:
1. Инициация.
2. Элонгация.
3. Терминация.
Инициация
Репликация начинается в нескольких точках молекулы ДНК. Это связано с тем, что размер генома типичной клетки млекопитающих составляет около 109 пар нуклеотидов (п.н.). В ДНК средней по величине хромосомы человека содержится несколько десятков миллионов п.н. При скорости репликации примерно 1000 пар нуклеотидов в минуту для полного завершения репликации молекулы ДНК в хромосоме понадобится несколько суток. Для клетки такой срок репликации генома совершенно не приемлем, т.к. во время репликации ДНК не осуществляется важный процесс – транскрипция РНК. Во время репликации ДНК практически все гены клетки, контролирующие её метаболизм, жизнедеятельность, адаптацию и специфические клеточные реакции выключены. Это связано с тем, что два процесса – синтез ДНК (репликация) и РНК (транскрипция) с их громоздким ферментативным обеспечением – одновременно на одной и той же матрице (материнской ДНК) происходить не могут. Поэтому в момент репликации происходит почти полное выключение транскрипции. Это ставит клетку в неблагоприятные условия, т.к. отсутствие экспрессии генов приводит к недостатку белков-ферментов, и, следовательно, к нарушению нормального течения метаболизма. Однако следует отметить, что природа предусмотрела специальные механизмы, которые снижают для клетки опасность выключения транскрипции. Например, в период предсинтеза ДНК (т.е. перед началом репликации) в цитоплазме клетки заранее накапливается достаточное количество всех видов РНК для поддержания синтеза белка в период репликации ДНК на необходимом для жизнедеятельности клетки уровне. Поэтому во время синтеза ДНК трансляция осуществляется (хотя и не в полном объёме), транскрипция же практически не идёт. В любом случае ещё раз подчеркнём, что время, на протяжении которого идёт процесс репликации ДНК, для клетки очень опасно. Поэтому не случайно в эво-
люции были созданы механизмы, до предела сокращающие время репликации ДНК в хромосомах. Таких механизмов несколько, но один из них достаточно прост: репликация начинается одновременно в нескольких точках молекулы ДНК и идёт от этих точек в разных направлениях. Приблизительные расчёты показали, что в молекуле ДНК одной хромосомы таких точек насчитывается несколько тысяч.
Такое большое количество мест, в которых начинается репликация ДНК хромосомы, несомненно существенно сокращает время её синтеза. Точки, в которых начинается репликация, носят название точки начала репликации (т.н.р.) или ориджин, ori-сайт(от англ. origin – начало). Эти точки представляют собой небольшие участки ДНК с определённой специфической последовательностью нуклеотидов. Начальным моментом репликации является присоединение к ори-сайтам специальных белков – инициаторных белков.
Рассмотрим два положения, касающихся ori-сайтов. Во-первых, репликация начинается не со всех точек одновременно. В одних точках она может пройти значительный участок, а в других только начаться. Во-вторых, от ori-сайтов процесс синтеза ДНК может двигаться в одном направлении или сразу в обоих противоположных направлениях – двунаправленная репликация (рис. 2). У эукариот чаще всего наблюдается второй тип репликации.
Наличие ori-сайтов привело к созданию понятия репликон.Репликон - это фрагмент матричной (материнской) ДНК, на котором идёт автономный синтез дочерней ДНК. Он начинается с точки начала репликации и заканчивается точкой, где фермент заканчивает репликацию.
 Е
Е
 |









 А Г Д ДНК
А Г Д ДНК
Б В
 Рис. 2. Схема расположения точек начала репликации. Стрелками обозначено направление синтеза ДНК от т.н.р.
Рис. 2. Схема расположения точек начала репликации. Стрелками обозначено направление синтеза ДНК от т.н.р.
У репликона три основных свойства:
а) он автономен;
б) обеспечивает процесс синтеза ДНК;
в) имеет собственное ферментативное обеспечение.
Если репликация однонаправленна, то протяжённость репликона соответствует промежутку между ori-сайтами, например репликон Б – В. Если же репликация двунаправленна (например между ori-сайтами Г – Д), то между этими точками будут располагаться два репликона Г – Е и Д – Е, где точка Е – конец двух различных процессов репликации, имеющих собственное ферментативное обеспечение. Как показали эксперименты, определить среднюю протяжённость одного репликона достаточно трудно. В связи с этим чаще всего за репликон принимают последовательность ДНК, ограниченную двумя ori-сайтами.
В точке начала репликации происходят события, которые связаны с присо-
единением к инициаторному белку других ферментов, запускающих целый ряд подготовительных процессов. В соответствии с очерёдностью эти события располагаются в следующем порядке:
а. Образование репликационной вилки.
б. Образование РНК-затравки.
а. Образование репликационной вилки
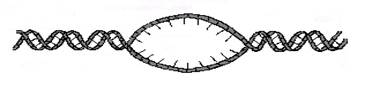
Образование репликационной вилки происходит в точках начала репликации. Вилка репликации представляет собой отошедшие друг от друга свободные нити матричной ДНК, на которых происходит синтез дочерних нитей ДНК. Схема образования вилок репликации представлена на рисунке 3. Две вилки репликации образуют глазок репликации. В обеих вилках репликации происходит синтез ДНК в противоположных направлениях. В глазке репликации вилки двигаются, расплетая ДНК, в противоположных направлениях.
Образованию вилок репликации предшествует целый ряд подготовительных процессов, которые, как правило, располагаются в следующем порядке:
· Освобождение молекулы ДНК от связи с гистонами в нуклеосоме.
· Деспирализация ДНК.
· Разрыв водородных связей между нитями матричной ДНК.
· Расхождение нитей ДНК.
· Фиксация матричных нитей ДНК.
Освобождение молекулы ДНК от связи с гистонами необходимый элемент периода инициации. В ядре клеток ДНК практически никогда не находится в свободном состоянии. Она связана с гистоновыми белками тетрамера и гистоном Н1. Кроме того, с ДНК взаимодействуют различные негистоновые белки, двухвалентные металлы, липиды и др. соединения. Всё это приводит к сильной спирализации отдельных участков ДНК в предсинтетическом периоде.
Каким образом происходит освобождение ДНК от связей с окружающими молекулами до конца не ясно. Однако известно, что в основе этого процесса лежат несколько химических реакций, осуществляемых ферментами – метилирование, ацетилирование и фосфорилирование положительно заряженных групп гистоновых и негистоновых белков. В результате белки теряют положительный заряд, их связи с отрицательно заряженной молекулой ДНК становятся слабыми. Это способствует высвобождению ДНК.
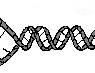
Сразу же после освобождения от связей с белком (или параллельно с этим) происходит процесс деспирализации ДНК. Этот процесс необходим для последующего освобождения нитей ДНК от связи друг с другом. Важное значение имеет и другой процесс. Нити молекулы ДНК не просто лежат параллельно друг другу, они взаимно закручены (см. рис. 4).
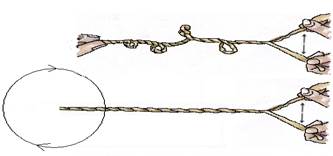
Существует множество способов раскрутки спиралей. Наиболее часто употребляемые способы показаны на рисунке 5. Очевидно, что способ А не совсем удобен, т.к. при фиксации одного конца и разведении противоположных концов создаются дополнительные силы, приводящие к формированию вторичных суперспиралей. При втором способе (противоположный конец свободен)
раскручивание приведёт к вращению свободного конца, а это может превратить ядро в субмолекулярный миксер, что небезопасно для клетки.
Ori-сайт
 |

 ДНК ●
ДНК ●

ДНК
 |

ДНК 

Репликативный глазок (состоит из двух репликационных вилок)
 |  |
ДНК
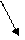 |  | ||
Левая вилка Правая вилка
Две репликационные вилки
 Рис. 3. Схема образования репликационных вилок в точке начала репликации. Пунктирными стрелками показано направление движения вилок
Рис. 3. Схема образования репликационных вилок в точке начала репликации. Пунктирными стрелками показано направление движения вилок
Рис. 4. Схема взаимного расположения нитей ДНК. Водородные связи между нитями не показаны
Для их раскручивания используются ферменты, которые осуществляют разрезание, вращение и воссоединение нитей.
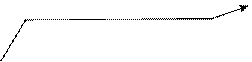
Принцип раскручивания спирали ДНК до конца не ясен. Один из возможных вариантов показан на рисунке 6. Суть его в том, что специальный фермент производит временный одноцепочечный разрыв в одной нити ДНК. И через этот разрез проходит (протаскивается) противоположная нить ДНК. При этом один виток спирали распрямляется. Затем разрез ликвидируется. Процесс
разрезания, расплетения и сшивки может повторяться множество раз по мере продвижения репликационной вилки по молекуле ДНК.
Все реакции, связанные с раскручиванием нитей ДНК осуществляют ферменты топоизомеразы. Уместно отметить, что аналогичным образом действуют эти ферменты и при другом процессе – транскрипции. Следует отметить, что одновременно специальные ферменты хеликазы разрывают водородные связи между оппозитными (противоположными) нитями ДНК делая их свободными.
А
Б
Рис. 5. Способы раскручивания верёвки. А – противоположный конец фиксирован, Б – противоположный конец свободен
Освобождённые из дуплекса нити ДНК долго оставаться в свободном состоянии не могут. Они или вновь соединяются, или взаимодействуют с активными реагентами из окружающего пространства, или формируют шпильки. Любой вариант способен нарушить нормальный ход репликации. Для предотвращения этого существуют специальные белки –SSB-белки. Эти белки обладают сродством к одноцепочечной ДНК и с появлением последней сразу же соединяются с ней на всём её протяжении, составляя жёсткий каркас, препятствуя не только её реакциям, но и образованию шпилек (рис. 7, 1).
Важным свойством этих белков является то, что они оставляют открытыми для реакций нуклеиновые основания одноцепочечной нити ДНК, что чрезвычайно важно для последующего синтеза дочерних нитей ДНК.
б.Образование РНК-затравки
После завершения всех вышеперечисленных процессов репликативная вилка почти готова к синтезу ДНК и выглядит как показано на рисунке 7, 1. Однако для полной готовности ДНК к репликации не хватает одной существенной детали. Синтез дочерних нитей в соответствии с матричными свободными нитями ДНК осуществляет фермент ДНК-полимераза. Однако начать процесс синтеза новой нити этот фермент не может. Он способен только продолжить синтез уже начатой цепочки. Поэтому другой фермент ДНК-зависимая РНК-полимераза (РНК-праймаза) вначале синтезирует на обеих разошедшихся в репликационной вилке матричных нитях ДНК небольшой участок РНК. Эта РНК носит название РНК-затравка (РНК-праймер или просто праймер) (на рис. 7 они обозначены буквами «А» и «Б»). РНК-затравки синтезируются параллельно матричным нитям ДНК по принципу комплементарности. Причём на одной нити РНК-затравка
(на рис. 7 она обозначена как «А») синтезируется, начиная с ori-сайта в сторону вилки репликации, на оппозитной нити РНК-затравка (на рис. 7 она обозначена как «Б») синтезируется, отступя от ori-сайта в его сторону.
Разрез в нити № 2 и протягивание через разрез нити № 1

 1
1
А


 Б 2
Б 2
В
Г
Д
Рис. 6. Схема деспирализации отрезка молекулы ДНК путём разрезания одной нити, протаскивания другой, раскручивания и сшивки разреза после деспирализации участка. Водородные связи между нитями не показаны
Синтезом двух РНК-затравок на двух противоположных нитях матричной ДНК заканчивается период инициации.
Элонгация
Фермент ДНК-полимераза начинает синтез новых нитей ДНК с 3’-концов двух РНК-затравок (А и Б на рис. 7, п. 2). При таком синтезе дочерние молекулы ДНК на матричных нитях будут синтезироваться в противоположных направлениях.
Элонгация начинается с присоединения одного нуклеотида к 3’-концу РНК-затравки. Это присоединение осуществляет фермент ДНК-полимераза.К первому нуклеотидуДНК-полимераза присоединяет второй, третий и т.д. нуклеотид в соответствии с принципом комплементарности к нуклеотидам, находящимся на матричной нити ДНК. Основания нуклеотидов новой нити и матричной соединяются друг с другом водородными связями. Поскольку синтез новой нити ДНК начинается с 3’-конца РНК-затравки, к которому присоединяется 5’-конецпервого нуклеотида ДНК, то принято говорить, что синтез дочерней нити ДНК происходит в направлении 5’– 3’. На той матричной нити, где синтез
дочерней нити идёт в сторону вилки репликации, он идёт непрерывно, и по мере раскручивания нити фрагмент синтезированной ДНК будет постоянно удлиняться. Чем дальше продвигается вилка репликации, тем длиннее будет вновь синтезированная цепь ДНК. Эту нить ДНК называют непрерывной, лидирующей или ведущей. В дальнейшем никаких РНК-затравок на матричной нити ДНК, с которой реплицируется лидирующая нить, не формируется. Сложнее происходит синтез дочерней ДНК на противоположной нити материнской ДНК, которая имеет направление 5’- 3’. К синтезированной на этой нити РНК-затравке (на рис.7 обозначена как «Б») нуклеотиды будут присоединяться также к 3’-концу и новая цепь будет синтезироваться в сторону от вилки репликации (рис.7, 2). Эту нить дочерней ДНК называют отстающей, запаздывающей. Таким образом, в репликативной вилке одновременно синтезируются две дочерние нити ДНК – ведущая (она синтезируется в направлении движения вилки репликации) и отстающая (синтезируется в направлении противоположном движению вилки репликации), т.е. направление синтеза обеих дочерних цепей противоположно.
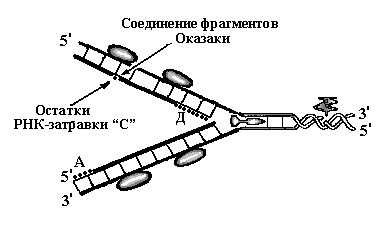
Синтез запаздывающей нити ДНК происходит не постоянно, а фрагментами. После окончания синтеза одного фрагмента вблизи репликационной вилки вновь происходит синтез РНК-затравки «С». С 3’-конца этой затравки вновь начинается синтез дочерней нити ДНК в направлении РНК-затравки «Б» и по её достижению синтез вновь прекращается – сформировался второй фрагмент ДНК, который начинается от РНК-затравки «С». Фактически на отстающей цепи мы имеем два фрагмента, которые состоят из соединённых друг с другом РНК и ДНК. Один фрагмент ДНК начинается с РНК-затравки «Б», второй – с РНК-затравки «С». Эти фрагменты ДНК носят название фрагменты Оказаки(рис. 7,3).
Последующие раунды репликации повторяются – топоизомераза раскручивает очередной виток спирали ДНК, хеликаза разрывает водородные связи между нитями ДНК и они расходятся, белки SSB фиксируют нити, лидирующая цепь продолжает удлиняться, на отстающей цепи синтезируется третий фрагмент РНК-затравки – «Д» и с его 3’-конца начинает синтезироваться новый фрагмент Оказаки. Затем на отстающей нити ДНК РНК-затравки разрушаются и оставшиеся фрагменты ДНК соединяются в единую цепь (рис. 8). Таким образом, на отстающей нити идут непрерывно 4 процесса: образование новых РНК-затравок, синтез с их 3’-конца фрагментов Оказаки, разрушение РНК-затравок и воссоединение фрагментов в единую цепь.
Терминация
Синтез ДНК продолжается до тех пор, пока не встретятся две репликативные вилки или когда репликативная вилка не подойдёт к концу молекулы ДНК в хромосоме. После встречи репликативных вилок синтезированные дочерние цепи ДНК из соседних вилок соединяются ферментом. Сложнее происходит терминация в случае, если репликативная вилка подошла к концу молекулы ДНК. Но и здесь репликацию прекращают специальные ферменты (рис. 9).
3. Коррекция (контроль) репликации
При репликации ДНК происходит важный процесс коррекции вставляемых нуклеотидов (рис. 10). В область синтеза ДНК поступают все четыре нуклеотида (на рис. 10 они обозначены как «претенденты нуклеотиды»). ДНК-полимераза отбирает нужного предшественника путём пробного спаривания его с нуклеотидом ДНК-матрицы.
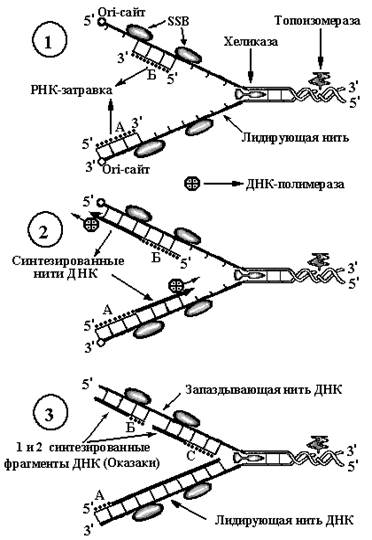
Рис. 7. Схемы репликации ДНК
 Правильность подобранного нуклеотида оценивается по нескольким параметрам, два из которых наиболее существенны. Во-первых, подобранный нуклеотид должен формировать столько же водородных связей, сколько и нуклеотид на матричной нити.
Правильность подобранного нуклеотида оценивается по нескольким параметрам, два из которых наиболее существенны. Во-первых, подобранный нуклеотид должен формировать столько же водородных связей, сколько и нуклеотид на матричной нити.
Рис. 8. Схема репликации ДНК – стадия элонгации. На запаздывающей нити синтезировался третий участок РНК-затравки – «Д». РНК-затравки «Б» и «С» разрушены и фрагменты Оказаки, синтезированные с их концов соединены в единую нить. На лидирующей цепочке синтез идёт непрерывно, РНК-затравка «А» разрушается
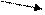

 Ориджины Лидирующая цепь
Ориджины Лидирующая цепь
 | |||
 | |||
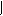









 3’ 5’
3’ 5’










 5’ 3’
5’ 3’
 | |||||
 | |||||
 | |||||

 Отстающая цепь
Отстающая цепь
 Репликон
Репликон

 Движение вилок Движение вилок
Движение вилок Движение вилок
Рис. 9. Схема осуществления синтеза ДНК в двух вилках репликации
Во-вторых, расстояние между сахарофосфатными остовами должно быть постоянным и соответствовать трём гетероциклическим кольцам в двух основаниях (одно кольцо, соответствующее пиримидиновому основанию в одной нити, и два кольца, соответствующие пуриновому основанию в другой нити). Если же претендент нуклеотид и нуклеотид в матрице будут содержать по два кольца в молекуле или по одному, то соединение не происходит.
Проверка правильного включения нуклеотида в растущую цепь производится дважды: один раз перед включением его в состав растущей цепи и второй раз перед включением следующего нуклеотида.
После того как прошла проверка на совместимость нуклеиновых оснований, формируется связь в сахарофосфатном остове.
4. Конвариантная редупликация как основа мутационной изменчивости
Этот термин ввёл в научный обиход учёный-генетик Тимофеев-Ресовский. Дословно конвариантная редупликация означает самовоспроизведение с изменением. Этот процесс лежит в основе всего эволюционного процесса.
Несмотря на высокую точность процессов репликации и эффективно работающую систему коррекции, во вновь синтезированных нитях ДНК всегда имеются нарушения. Эти дефекты чаще всего носят название «генные мутации». Подсчёты показали, что они происходят с частотой 1 ошибка на 1010 пар нуклеотидов. Редупликация, происходящая с ошибками, носит название конвариантная редупликация.
В основе нарушений лежат самые различные причины. Например, высокая или наоборот очень низкая концентрация нуклеотидов в месте синтеза, спонтанная потеря пуриновых оснований (см. далее), дезаминирование цитозина, который превращается при этом в урацил под действием ультрафиолетового облучения, присутствие в месте синтеза химических мутагенов и т.д. Судьба возникших ошибок неоднозначна. Они могут быть исправлены репарационными процессами (см. далее). Это наиболее благоприятный для клетки вариант. В некоторых случаях повреждение в молекуле ДНК не устраняется. При этом возможны два варианта разворачивания событий. Если повреждение молекулы ДНК затронуло функционально не активную область ДНК, то фенотипически такая ошибка не будет выявляться и, как правило, никаких тяжёлых последствий наблюдаться не будет. Иное дело, если ошибка спаривания произошла в нуклеотидных последовательностях какого-либо гена. В этом случае вероятность появления фенотипических нарушений увеличивается. Это может привести к гибели клетки или целого организма. При этом вместе с погибшей клеткой элиминируется и дефект ДНК. И, наконец, следует учитывать, что генные мутации лежат в основе мутационной изменчивости. А она приводит к возникновению множественных аллелей, которые делают генофонд популяций более пластичным.
5. Метилирование ДНК
И в заключении необходимо остановиться на важном процессе, который происходит в момент или сразу же после синтеза дочерних цепей ДНК – метилирование вновь синтезированных цепей. У человека специальный фермент ДНК-метилаза метилирует цитозин, присоединяя к нему группы СН3. Процесс метилирования скорее всего необходим для формирования структуры хромосом (упаковки ДНК) и регуляции транскрипции генов. Кроме того, метилирование спо-собствует инактивации одной Х-хромосомы у млекопитающих. У бактерий метилирование защищает ДНК от разрезания своими собственными ферментами.
Сахарофосфатный остов
 |  |
Водородные


 связи
связи
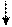
 Т А
Т А



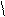


 Г Ц
Г Ц
 | 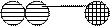 | | |||


 Дочерняя цепь
Дочерняя цепь
 ДНК
ДНК

 А Т
А Т
 |  |  | |||


 Ц
Ц






 А
А




 Г
Г
 | |||||||||
 | |||||||||
 | |||||||||
 | |||||||||
 | |||||||||
Ц


 Матричная цепь ДНК Т
Матричная цепь ДНК Т
 | |  | ||||||
 | ||||||||
 | ||||||||
Претенденты нуклеотиды
Рис. 10. Схема подбора комплементарного основания к цитозиновому нуклеотиду (Ц). Среди претендентов с этим нуклеотидом совместим только гуаниновый нуклеотид (Г), т.к. он формирует три водородные связи и имеет два кольца
6. Лекарственные препараты и репликация
Процесс репликации подвержен воздействию самых различных факторов и, в частности, лекарственных препаратов. Так, например, дауномицин и другие противоопухолевые препараты имеют в своей молекуле плоскую циклическую структуру, которая встраивается (интеркалируется) между парами оснований. Это ведёт к локальному изменению структуры ДНК, в результате чего ферменты репликации прекращают свою работу. Например, топоизомераза теряет способность деспирализовывать ДНК в месте интеркаляции. Такие алкилирующие
вещества, как тиофосфамид и др., модифицируют основания путём присоединения к ним алкильных группировок. Если ферменты репликации встречают это основание, их работа может прекратиться.
Ингибитор ДНК-топоизомеразы – новобиоцин, вмешиваясь в работу фермента, прекращает деспирализацию ДНК, а, следовательно, и синтез РНК.
Резюме. Репликация ДНК – процесс точной передачи информации от ДНК на ДНК в результате самовоспроизведения матричных молекул. В репликации различают три периода.
1. Инициация. Происходит образование репликационной вилки и образование РНК-затравки. Синтез начинается одновременно на множестве участков ДНК. Перед синтезом ДНК деспирализуется, водородные связи разрываются и нити отходят друг от друга.
2. Элонгация. Синтез РНК начинается с РНК-затравки и идёт одновременно на обеих нитях материнской ДНК. На одной нити синтез идёт непрерывно, на другой – фрагментами (фрагменты Оказаки).
3. Терминация. Синтез РНК заканчивается при встрече репликационных вилок или на конце молекулы ДНК.
| РЕПАРАЦИЯ ДНК |
1.Общее представление о репарации
Процесс репарации характерен для всех живых существ. Это одно из основных свойств наследственного материала, напрямую связанное с интенсивностью процессов старения, динамикой развития опухолевых заболеваний, формированием врождённых пороков развития, возникновением наследственной патологии и др. Изменения ДНК, не подвергшиеся репарации, сыграли (и играют) важную роль в эволюционном формировании новых адаптивных свойств живого.
Остановимся на трёх важных принципах, характерных для процесса репарации.
1. Ферменты репарации определяют место большинства повреждений на молекуле ДНК по её локальной денатурации в месте повреждения.
Эффективность любой репарационной системы, в первую очередь, зависит от того насколько быстро и точно ферменты обнаружат места повреждений ДНК. Поскольку характер последних чрезвычайно разнообразен, то трудно представить существование многочисленных ферментов, опознающих «свои» специфические повреждения ДНК. В действительности всё оказалось проще. Любое нарушение первичной структуры ДНК приводит к одинаковым последствиям – сразу же возникает нарушение в структуре второго, третьего и т.д. порядка, т.е. в месте повреждения наступает локальная денатурация ДНК. Возникшие в ДНК участки денатурации (в отличие от первичных повреждений) не специфичны. Это позволяет специальным ферментам быстро выявлять денатурированные
участки ДНК и подключать для их восстановления специфические репарационные системы.
2. Повреждённый участок на одной нити ДНК восстанавливается в соответствии с неповреждённым оппозитным участком второй нити. При повреждении двух нитей в восстановлении повреждения участвуют сестринские нити хроматид.
Отмечена закономерность, что при воздействии на ДНК мутагенных факторов, повреждения в основном касаются одной нити ДНК. Одновременное повреждение обеих нитей случается не часто и они, как правило, заканчиваются гибелью клетки. Восстановить повреждённый участок не трудно, если имеется план его первоначального (до повреждения) строения. Такой план всегда имеется – это последовательность нуклеотидов на второй, оппозитной цепочке ДНК, т.е. в нити ДНК, противоположной месту повреждения. Здесь располагаются нуклеотиды, комплементарно отражающие последовательность нуклеотидов повреждённого участка. Поэтому многие репарационные системы «ремонтируют» повреждённый участок, застраивая его комплементарными нуклеотидами в соответствии с планом оппозитного участка.
Иное дело, если повреждаются одновременно обе нити ДНК. Это бывает, когда молекула ДНК с повреждением вступает в период синтеза ДНК. В этом случае на дефектной ДНК происходит синтез дочерних нитей ДНК, причём на повреждённой нити синтезируется дефектная дочерняя нить. Но и в этом случае план повреждённых участков сохраняется в молекуле ДНК, которая сформировалась в результате репликации неповреждённой нити ДНК. При максимальном сближении сестринских хроматид в профазе мейоза и митоза происходит репарация повреждённых нитей ДНК в соответствии с неповреждёнными участками молекулы ДНК сестринской хроматиды.
У диплоидных организмов при репарации повреждённого участка ДНК возможно использование плана строения неповреждённого участка ДНК гомологичной хромосомы.
3. Энергия на процесс репарации черпается в основном из АТФ. Но в некоторых случаях используется энергия фотона света, который активирует ферменты репарации.
Репарация ДНК – энергозависимый процесс. В качестве поставщика энергии используются макроэргические соединения (например, АТФ). Однако есть одно исключение – это фотореактивация (световая репарация). Фермент, осуществляющий эту реакцию, использует энергию солнечного света, а не аккумулированную в химических связях АТФ. Называется этот фермент – фотолиаза. Фермент репарирует повреждения, связанные с образованием тиминовых димеров (см. далее).
2.Виды репарации
Прежде всего, все виды репарации (или реакции) делятся на две большие группы – прямые и непрямые. Прямые реакции отличаются простотой и непо-
 Сахарофосфатный остов
Сахарофосфатный остов


Водородные


 связи
связи

 Г Ц
Г Ц





 | |||
 |


 УФ Т А
УФ Т А
 | |||
 | |||
 Т А
Т А
 | |||
 |
 Г Ц
Г Ц
 | |||
 | |||

Рис. 11. Схема образования тиминовых димеров под действием УФ-облучения. Образовавшиеся химические связи между тиминами одной нити показаны пунктирными линиями
средственным действием на повреждённый объект. Никаких «предварительных работ» не проводится. Практически все они функционируют до периода синтеза ДНК. К ним относится фотореактивация. При непрямом типе репарации имеющееся повреждение ДНК вначале вырезается. В результате на одной из нитей формируется брешь, которая застраивается специальными ферментами в соответствии с планом расположения нуклеотидов на противоположной цепи. К этому типу репарации относится эксцизионная репарация и ряд других.
Прямой тип репарации
Фотореактивация
Неоспоримым фактом является то, что ультрафиолетовый свет (УФ) повреждает уникальную молекулу ДНК. Повреждения в основном касаются пиримидиновых оснований – тимина и цитозина. Под влиянием УФ света происходит
перераспределение валентностей в пиримидиновых основаниях, т.е. между нахо-
дящимися рядом на одной нити двумя тиминами, двумя цитозинами или тимином и цитозином. Следствием этого является появление необычной химической связи между основаниями и возникновение димера (два нуклеотида) – тиминового (тимин-тимин), цитозинового (цитозин-цитозин) или тимин-цитозинового. Водородная связь между оппозитными нуклеотидами при этом разрушается (рис.11). Есть данные о том, что пиримидиновые димеры способны активировать процессы, приводящие к развитию опухолевых заболеваний.
Возникшая мутация репарируется несколькими системами репарации. Одна из них фотореактивация. Основной фермент этой реакции белок фотолиазаимеет сложную белковую структуру. На одном участке молекулы находится светочувствительный центр, воспринимающий фотоны синего света и активирующий фермент. Фермент в таком состоянии находит димеры в молекуле ДНК, разрывает образованные УФ связи между тиминами и восстанавливает межнитевые водородные связи пиримидин – пурин. По завершению цикла фермент отходит от ДНК.
Следует отметить, что при этом типе репарации фермент непосредственно действует на повреждение, восстанавливая его. Это классический тип прямой репарации.
Непрямой тип репарации
Эксцизионная репарация
Надрез Расширение бреши Застройка бреши Сшивка

 (Эндонуклеаза) (Экзонуклеаза) (ДНК-полимераза) (Лигаза)
(Эндонуклеаза) (Экзонуклеаза) (ДНК-полимераза) (Лигаза)
 | |||||
 | |||||
 |

Тиминовый димер
Рис. 12. Схема эксцизионной репарации тиминовых димеров
Тиминовые димеры могут быть в силу ряда причин не восстановлены системой фотореактивации. В этом случае активируются ферменты системы эксцизионной репарации. Этот тип репарации позволяет исправить до 95% спонтанных мутаций. Существует несколько типов реакций эксцизионной репарации. Их объединяет то, что повреждённое основание, нуклеотид или неправильно спаренное основание (мисмэтч) вначале полностью вырезается из цепи, а затем вставляется новый нуклеотид (или нуклеотиды), комплементарный нуклеотиду противоположной цепочки. Приведём в качестве примера эксцизионную репара-
цию нуклеотидов(тиминовых димеров). Реакция состоит из нескольких этапов. На первом этапе фермент эндонуклеаза находит повреждённую нить ДНК и делает надрез в месте повреждения. Другой фермент, экзонуклеаза, расширяет надрез, «откусывая» с обоих концов надреза нуклеотиды вместе с тиминовым димером. Образовавшаяся брешь застраивается с одного конца нити ДНК специальным ферментом ДНК-полимеразой. При этом вставляемые нуклеотиды комплементарны нуклеотидам неповреждённой нити ДНК. Незастроенным остаётся небольшой разрыв, который сшивается ДНК-лигазой (рис.12).
ОБЩЕЕ ПРЕДСТАВЛЕНИЕ О ТРАНСКРИПЦИИ
Экспрессия гена – это реализация генетической информации, закодированной в ДНК, путём её транскрипции и трансляции. Транскрипция – первый этап экспрессии генов. Заключается она в переводе информации, содержащейся в гене, на РНК путём синтеза последней на одной нити ДНК гена. В результате транскрипции синтезируются все виды РНК – информационная (иРНК), рибосомальная (рРНК), транспортная (тРНК) и другие (регуляторные, малые ядерные и пр.). Все они принимают участие в экспрессии генов, но только иРНК переносит информацию о строении белка с нуклеотидного текста ДНК на аминокислотный текст белка. Все остальные виды РНК обеспечивают эффективное осуществление этого процесса.
Суть транскрипции заключается в следующем: специальные ферменты подготавливают молекулу ДНК в области гена к транскрипции (раскручивают спираль ДНК, разрывают водородные связи между нитями и т.д.). Фермент ДНК-зависимая РНК-полимераза синтезирует РНК на матричной нити ДНК от стартовой точки до точки окончания транскрипции. Синтез РНК заключается в последовательном наращивании в ней нуклеотидов, комплементарных матричной нити ДНК. Исключение составляет урацил РНК, который вставляется против аденина матричной нити ДНК.
В транскрипции принимает участие множество ферментов, но непосредственно синтез РНК осуществляет фермент ДНК-зависимая РНК-полимераза (или просто РНК-полимераза). У прокариот все виды РНК (иРНК, рРНК, тРНК) синтезируются одной РНК-полимеразой, а у эукариот они синтезируются тремя разными ферментами: РНК-полимеразой I, РНК-полимеразой II и РНК-полимеразой III.
Информационные РНК у эукариот транскрибируются РНК-полимеразой II.
Транскрипция – это сложный многоэтапный процесс и одна РНК-полимераза не в состоянии полностью обеспечить его. На разных этапах транскрипции к РНК-полимеразе присоединяются и, наоборот, отщепляются различные белковые субъединицы, которые модифицируют её активность в соответствии с требованиями данного этапа.
Транскрипция, как и все процессы матричного синтеза у про- и эукариот, состоит из трёх этапов – инициации, элонгации и терминации. В дальнейшем процесс транскрипции будем рассматривать только у эукариот.
1. Периоды транскрипции
Инициация
Инициация – это подготовительный этап. В эту стадию происходит множество синхронизированных во времени процессов. Рассмотрим два из них:
1. Формирование инициаторного комплекса.
2. Образование «транскрипционного глазка».
РНК-полимераза
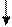 | |||
 | |||

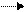
 ТФ ТФ
ТФ ТФ
 |  |  | |||
Специальный 
 белок
белок

Промотор
 |
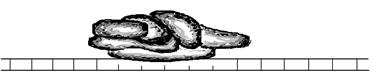 Инициаторный комплекс
Инициаторный комплекс
Промотор
Т А Т А
Рис. 13. Присоединение РНК-полимеразы и транскрипционных
факторов к ТАТА-боксу промотора у эукариот
Формирование инициаторного комплекса происходит на промоторе. Сама РНК-полимераза, как правило, не может связаться с промотором. Поэтому вначале специальный белок взаимодействует со специфической областью на промоторе. В этой области располагается определённая последовательность нуклеотидов. Эта последовательность носит название бокс Прибнова. Эта последовательность нуклеотидов богата адениновыми и тиминовыми нуклеотидами– ТАТА. Отсюда и другое название этого участка промотора

