 2015-05-13
2015-05-13 3413
3413Изучение сна, так же как и циркадианных ритмов, началось сравнительно недавно. Первые экспериментальные работы, посвященные мозговым механизмам сна, были проведены в 30-х годах бельгийским нейрофизиологом Бремером (Bremer), который производил перерезки ствола мозга на различных уровнях у кошек. Подобные операции осуществлялись уже давно с целью изучать подавление и облегчение спинальных рефлексов, однако Бремер в отличие от его предшественников заинтересовался влиянием таких перерезок на вышележащие отделы. Оказалось, что перерезки на уровне моста, продолговатого мозга и ниже не приводят к нарушению цикла сна—бодрствования у кошки, однако после более высоких перерезок (на уровне среднего мозга) наступал непрерывный сон. Это указывало на важную роль области среднего мозга и моста в поддержании бодрствования.
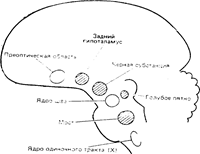
Рис.. Обобщенная схема мозговых структур, которые участвуют в регуляции сна и бодрствования. Центры бодрствования заштрихованы, центры сна изображены светлыми.
Исследования на человеке позволили вскрыть сложную внутреннюю структуру сна. Оказалось, что во время сна возникают периоды мозговой активности, в чем-то сходные с состоянием бодрствования. Таким образом, стало ясно, что сон—это не просто «отключение», что он, возможно, обусловлен взаимодействием нескольких нейронных систем. Поиск этих систем стал предметом многочисленных экспериментов на животных (главным образом на кошках), осуществленных в 60-х и 70-х годах. Например, французский исследователь Мишель Жувэ изучал последствия разрушения отдельных структур ствола мозга. В этих и других работах применялись методы выявления различных медиаторов, и изучалось действие их антагонистов. Кроме того, проводились опыты с раздражением специфических областей ствола мозга и базальных отделов переднего мозга, а также с регистрацией активности отдельных клеток этих структур.
«Центры бодрствования». Активацию и D-сон легче всего вызвать путем высокочастотного электрического раздражения ретикулярной формации ствола мозга (см. рис. 26.14), как показали, в частности, работы Моруцци и Мэгуна. Наиболее выраженный эффект возникает при стимуляции гигантоклеточных полей покрышки. Отростки нейронов этих полей широко ветвятся в пределах ствола. Вторым центром бодрствования служит голубое пятно. В нем находятся норадренэргические нейроны, отдающие многочисленные двигательные аксоны к большинству отделов переднего мозга, мозжечка и спинного мозга. Как показал Жувэ, разрушение голубого пятна у кошек приводит к избирательному выпадению стадий D-сна. Третьим «центром», ответственным за поддержание бодрствования является система дофаминэргических волокон, выходящих из черной субстанции и смежных участков среднего мозга. Повреждение этих волокон у кошек приводит к коматозному состоянию. Однако реакция активации ЭЭГ в ответ на сенсорные стимулы при этом сохраняется; кроме того, при повреждении тех же волокон у крыс не наблюдается аналогичных нарушений. Это указывает на то, что дофаминэргические пути, возможно, участвуют не столько в общей активации, сколько в «запуске» двигательной активности. Наконец, есть данные, что в механизмах бодрствования участвуют также базальные отделы переднего мозга. Активацию можно легко получить путем раздражения гипоталамуса, а удаление задних отделов гипоталамуса приводит к продолжительному сну (это впервые показал Валле Наута в Голландии в 1946 г.). Однако эти эффекты могут быть частично обусловлены волокнами срединного пучка переднего мозга, особенно теми, которые связывают ядра ствола с передним мозгом.
«Центры сна». Ряд исследований указывал на большое значение серотонинэргических нейронов ядер шва (в стволе мозга) для глубокого сна. Разрушение этих ядер у кошек приводит к полной бессоннице. Подавление синтеза серотонина ингибитором триптофангидроксилазы n-хлорфенилаланином тоже вызывает бессонницу, причем введение серотонина в какой-то степени восстанавливает сон. Вторым стволовым «центром сна» является ядро одиночного тракта, к которому подходят сенсорные волокна от вкусовых почек языка и от внутренних органов. Раздражение этого участка приводит к синхронизации ЭЭГ. С ядрами одиночного тракта связана так называемая area postrema. В этой области гематоэнцефалический барьер отсутствует, поэтому на нее могут влиять вещества, растворенные в крови; токсические агенты, вызывающие рвоту, воздействуют именно на клетки этой области. Серотонин здесь модулирует влияние ядра одиночного тракта на сон. Механизмы влияния этих двух отделов на сон и бодрствование неизвестны; возможно, что они обусловливают сонливость, возникающую после приема пищи, при некоторых обменных процессах и активности внутренних органов. Наконец, в регуляции сна и бодрствования участвует преоптическая область базальных отделов переднего мозга. Поражение этой области приводит к бессоннице, а ее электрическое раздражение—к синхронизации ЭЭГ 11 сонливости, переходящей в сон. В 1970 г. Бремер показал, что стимуляция преоптической зоны уменьшает степень активации, вызываемой раздражением активирующей ретикулярной системы.
1. Сон как системный процесс. Биологическое значение сна.
2. Объективные признаки сна.
3. Электоэнцефалографические показатели сна.
4. Медленноволновая и парадоксальная (быстроволновая) стадии сна. Электроэнцефалографический пародокс.
5. Корково-подкорковые механизмы сна.
6. Факторы, определяющие продолжительность сна.
7. Соматовегетативные проявления сна.
8. Фазы сна и психическая деятельность.
9. Механизмы формирования сновидений.
10. Теории сна.
11. Биологически активные вещества в механизмах сна.
12. Депривация сна.
Темы рефератов:
1. Нарушения сна.
2. Развитие стадии парадоксального сна в фило- и онтогенезе.
3. Механизмы формирования сновидений.
4. Память и сон.
5. Двигательная активность в период сна.
6. Развитие представлений о животном гипнозе.
7. Представления И.П.Павлова о переходных гипнотических фазах и сне.
8. Этапы формирования цикла бодрствование-сон.
9. Циркадные ритмы.
Литература:
1. Аладжалова Н.А., Кольцова А.В. Сверхмедленные ритмические колебания потенциалов мозга во сне //Функциональные состояния мозга. – М., 1975. – СС. 62-73.
2. Борбели А. Тайна сна. – М., 1981. – 191 с.
3. Динилин В.П., Малкин В.Б., Латаш Л.П. отражение особенностей биоэлектрической активности мозга бодрствующего человека в ЭЭГ сна // Физиология человека. – 1975. – Т. 1, № 6. – Сс. 1027-1030.
4. Данкелл С. Позы спящего. Ночной язык тела. – Нижний Новгород., 1991. – 239 с.
5. Латаш Л.П. Эмоции и сон // Физиология человека и животных. Т.15. Физиология эмоций. - М., 1975. – Сс. 94-152.
6. Латаш Л.П. Сон как сфера бессознательной активности // Бессознательное. Природа, функции, методы исследования. – Т.2. - Тбилиси, 1978.
7. Линдсли Д.Б. Внимание, сознание, сон, бодрствование //
8. Нейрофизиологические основы внимания. М., 1979.
9. Моисеева Н.И. Современные представления о механизмах регуляции и целевой функции сна //Успехи физиологических наук. – 1981. – Т.12., № 3. – Сс. 86-105.
10. Моисеева Н.И. Формирование цикла сон-бодрствование как эволюция биологической регуляции // Сон как фактор регуляции функционального состояния организма. – Л., 1985.
11. Психофизиология. Учебник для вузов /Под ред. Ю.И.Александрова. – СПб., 2001. – 496 с.

