 2015-05-26
2015-05-26 6762
6762«Для того чтобы понять жизнь растения... необходимо прежде ознакомиться с его формой: для того чтобы понять действие машины, нужно знать ее устройство».
К. А. Тимирязев
| 1.1 Строение растительной клетки |
| I.I.I Биологические мембраны и их функции |
Термин «клетка» (от греч. «cytos» — клетка или лат. «cellula» — полость) впервые применил Роберт Гук в 1665 г. при описании строения пробки, изученного с помощью усовершенствованного им микроскопа. С 1839 г., когда М. Я. Шлейденом и Т. Шванном была сформулирована клеточная теория, получила признание универсальность клеточного строения всего живого.
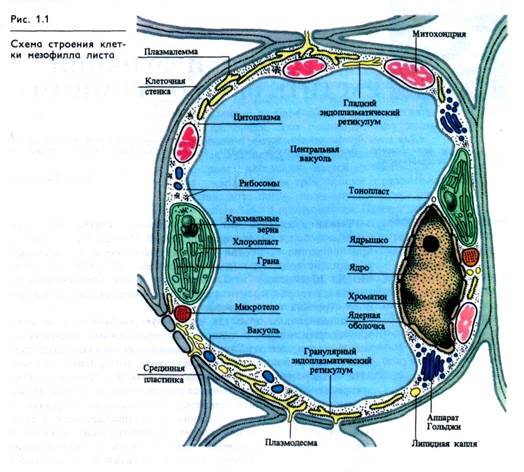
Растительная клетка как клетка эукариотического организма содержит ядро с одним или несколькими ядрышками, митохондрии, аппарат Гольджи, эндоплазматический ретикулум, микротела, рибосомы и полирибосомы, компоненты цитоске-лета — микротрубочки и микрофиламенты. В отличие от других эукариотических организмов для растительных клеток характерны: 1) пластидная система, возникающая в связи с фототрофным способом питания, 2) полисахаридная клеточная стенка, окружающая клетку, 3) центральная вакуоль в зрелых клетках, играющая важную роль в поддержании тургора (рис. 1.1). Кроме того, у делящейся растительной клетки нет центриолей. Электронно-микроскопические снимки свидетельствуют о том, что клеточная, или плазматическая, мембрана (плазмалемма) и внутриклеточные мембраны составляют основу ультраструктуры клеток эукариот.
Строение биологических мембран. В настоящее время наибольшим признанием пользуется жидкостно-мозаичная гипотеза строения биологических мембран. Согласно этой гипотезе основу мембраны составляет двойной слой фосфолипидов с некоторым количеством других липидов (галактолипидов, стеринов, жирных кислот и др.), причем липиды повернуты друг к другу своими гидрофобными концами. Ненасыщенные жирные кислоты полярных липидов обеспечивают несколько разрыхленное (жидкое) состояние бислоя при физиологических температурах. Этому же способствуют и стерины. Биологические мембраны уже по составу липидов построены асимметрично, так как две их стороны — наружная и внутренняя — обращены в качественно разные гидрофильные среды. В наружном слое плазмалеммы содержится больше стеринов и гликолипидов.
 Липиды, входящие в состав мембранного бислоя, не закреплены жестко, а непрерывно меняются местами. Перемещения липидных молекул бывают двух типов: 1) в пределах своего монослоя (латеральная диффузия) и 2) путем перестановки двух липидных молекул, противостоящих друг другу в двух монослоях («флип-флоп»). При латеральной диффузии молекулы липидов претерпевают миллионы перестановок в секунду, а скорость ее составляет 5—10 мкм/с. Перестановки молекул липидов из одного монослоя в другой происходят значительно реже, но могут ускоряться мембранными белками.
Липиды, входящие в состав мембранного бислоя, не закреплены жестко, а непрерывно меняются местами. Перемещения липидных молекул бывают двух типов: 1) в пределах своего монослоя (латеральная диффузия) и 2) путем перестановки двух липидных молекул, противостоящих друг другу в двух монослоях («флип-флоп»). При латеральной диффузии молекулы липидов претерпевают миллионы перестановок в секунду, а скорость ее составляет 5—10 мкм/с. Перестановки молекул липидов из одного монослоя в другой происходят значительно реже, но могут ускоряться мембранными белками.
В жидких слоях липидных мембран находятся специализированные протеиновые комплексы. Липопротеины погружены в липидную фазу и удерживаются гидрофобными связями (интегральные белки). Гидрофильные белки (периферические) удерживаются на внутренней и внешней поверхностях мембран электростатическими связями, взаимодействуя с гидрофильными головками полярных липидов. Основную роль в формировании мембран играют гидрофобные связи: липид — липид,
 |
липид — белок, белок — белок. Толщина биомембран не превышает 6—10 нм.
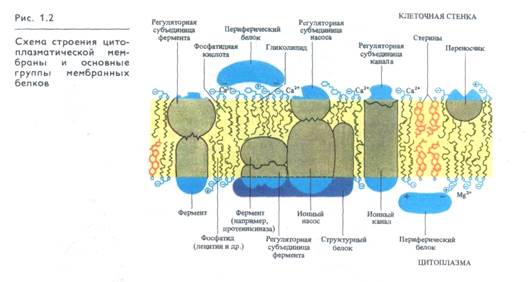
В состав мембран входят белки, выполняющие функции ферментов, насосов, переносчиков, ионных каналов, а также белки-регуляторы и структурные белки (рис. 1.2).
Интегральные белковые глобулы располагаются в фосфоли-пидных слоях мембран ориентированно. Эта ориентация определяется особенностями гидрофобной поверхности каждого белка, локализацией и свойствами его гидрофильных участков. Определенные участки липопротеиновых глобул (участки узнавания) служат для самосборки полиэнзимных мембранных комплексов. На положение белков в мембране оказывают влияние состав фосфолипидов, прочно связанных с глобулами, состояние «свободных» фосфолипидов двойного слоя, а также величина электростатического заряда мембраны. Функциональная активность мембран и изменения мембранного потенциала сопровождаются всплыванием или погружением субъединиц, их латеральными перемещениями. Предполагается, что такое перемещение белков в мембране может быть ограничено их связью с микрофиламентами и микротрубочками.
 Функции биологических мембран. Лабильная структура мембран позволяет выполнять им различные функции: барьерные, транспортные, осмотические, электрические, структурные, энергетические, биосинтетические, секреторные, пищеварительные, рецепторно-регуляторные и некоторые другие. Первичным назначением клеточной мембраны было отделение внутренней среды от внешней. Затем в процессе эволюции возникло большое количество специализированных внутриклеточных отсеков (компартментов), что позволило клетке и органоидам удерживать в небольших объемах необходимые ферменты и метаболиты, создавать гетерогенную физико-
Функции биологических мембран. Лабильная структура мембран позволяет выполнять им различные функции: барьерные, транспортные, осмотические, электрические, структурные, энергетические, биосинтетические, секреторные, пищеварительные, рецепторно-регуляторные и некоторые другие. Первичным назначением клеточной мембраны было отделение внутренней среды от внешней. Затем в процессе эволюции возникло большое количество специализированных внутриклеточных отсеков (компартментов), что позволило клетке и органоидам удерживать в небольших объемах необходимые ферменты и метаболиты, создавать гетерогенную физико-
химическую микросреду, осуществлять на разных сторонах мембраны разнообразные, иногда противоположно направленные биохимические реакции.
Но одновременно с появлением фосфолипидных мембранных барьеров должны были возникнуть и механизмы трансмембранного переноса ионов, субстратов, метаболитов, которые в ходе эволюции постоянно усложнялись. В живых клетках различают пассивный (по химическому и электрическому градиентам) и активный (против электрохимического градиента с затратой метаболической энергии) транспорт. Пассивный транспорт осуществляется: а) через фосфолипидную фазу, если вещество растворимо в липидах, б) по промежуткам между липидами, если такие промежутки появляются, в) с помощью липопротеиновых переносчиков, г) по специализированным каналам, образованным липопротеиновыми комплексами (натриевые, калиевые и другие каналы). Сахара, аминокислоты и некоторые другие субстраты могут переноситься специальными переносчиками в симпорте (т. е. совместно и в ту же сторону) с ионами Н+ (у бактерий, грибов и растений) или с Na+ (у животных), причем основной движущей силой в этом случае является градиент ионов, а не субстрата. Возможен также активный транспорт Сахаров в фосфорилированном и нефосфорилиро-ванном виде. Перенос ионов против электрохимического градиента, т. е. а ктивный транспорт осуществляется транспортными АТРазами с использованием энергии АТР. Известны К+, Na + -ATPaзa, Н + -АТРаза, Са2+-АТРаза, анионная АТРаза. Активный транспорт ионов Н+ может поддерживаться и за счет энергии NADH, NADPH или других окисляющихся соединений участками редокс-цепи. Перенос ионов Н+ через биологические мембраны с использованием АТР или NAD(P)H получил название протонной помпы (Н+- помпы, или Н+- насоса).
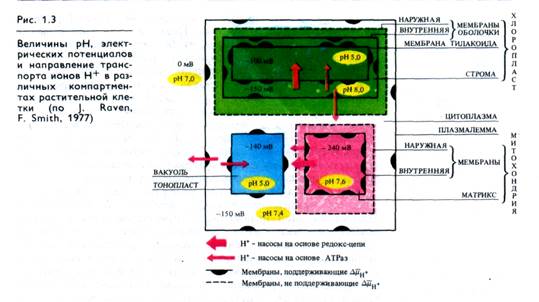
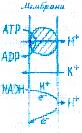 Н+- помпа так же важна для жизнедеятельности растительных клеток, как и Na +-помпа у животных. Она участвует в таких важнейших процессах (рис. 1.3), как регуляция внутриклеточного рН, создание мембранного потенциала, запасание и трансформация энергии, мембранный и дальний транспорт веществ, поглощение минеральных солей корнями, рост и двигательная активность и др.. При активации Н+- помпы мембранный электрохимический потенциал ионов Н+ (ΔµΗ+) возрастает и это возрастание включает в себя увеличение его электрического (ΔΨ) и химического (ΔрН) компонентов:
Н+- помпа так же важна для жизнедеятельности растительных клеток, как и Na +-помпа у животных. Она участвует в таких важнейших процессах (рис. 1.3), как регуляция внутриклеточного рН, создание мембранного потенциала, запасание и трансформация энергии, мембранный и дальний транспорт веществ, поглощение минеральных солей корнями, рост и двигательная активность и др.. При активации Н+- помпы мембранный электрохимический потенциал ионов Н+ (ΔµΗ+) возрастает и это возрастание включает в себя увеличение его электрического (ΔΨ) и химического (ΔрН) компонентов: 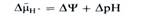
Оба компонента могут быть использованы на перемещение веществ. Электрический потенциал служит энергети-ческой основой для поглощения катионов (К +, Mg2 +, Mn2 +, Са2+ и др.), а протонный — для поступления в клетку анионов, Сахаров, аминокислот в симпорте с ионами Н+, а также для осморегуляции.
Многообразна роль осмотической работы, выполняемой биомембранами. Например, клетки пресноводных простейших и некоторых водорослей (эвгленовые, вольвоксовые), чтобы не быть разорванными, вынуждены постоянно сопротивляться пассивному осмотическому поступлению воды в клетки путем выведения избытка воды сократительными вакуолями. Для всех остальных растений осмотическое поступление воды не приводит к разрыву клеток, так как гидростатическому (тургорному) давлению противостоит противодавление эластически растянутых клеточных стенок. Более того, интенсивный напор воды становится важнейшим фактором существования растений. Тургор позволяет поддерживать форму неодревесневшим частям и является той силой, которая обеспечивает рост растяжением растительных клеток. Наконец, регулируя концентрацию осмотически активных веществ, растительные клетки всасывают воду из окружающей среды, даже если содержание воды в среде низкое. О мембранном транспорте растворенных веществ, который также входит в понятие осмотической работы, уже говорилось.
Структурная функция мембран заключается в упорядоченности расположения полиферментных комплексов, высокая эффективность которых определяется сближением каталитических центров. В среднем с одной молекулой погруженного в мембрану фермента контактируют 20 — 30 молекул фосфолипидов. Вследствие непрерывного движения липидов, примыкающих к белку, в липидном окружении фермента возникают молекулярные «дырки». Это явление не влияет на работу фермента, если липиды находятся в жидком агрегатном состоянии. При локальном «затвердевании» липидов возможно их устранение с достаточно больших участков белка, что может приводить к слипанию белков-ферментов и нарушению их функций. Поэтому для деятельности мембранных ферментов важно, чтобы окружающие их липиды находились в жидком агрегатном состоянии.
 Непроницаемость фосфолипидного бислоя для большинства соединений создает условия для векторных процессов. Векторный характер имеют транспорт веществ и передача сигналов из внешней среды в цитоплазму. Само расположение ферментов в липидной фазе мембраны весьма существенно для их активности. Таким образом, благодаря упорядоченности и векторности ферментов и транспортных механизмов осуществляется мембранная координация множества биохимических реакций, протекающих в клетке.
Непроницаемость фосфолипидного бислоя для большинства соединений создает условия для векторных процессов. Векторный характер имеют транспорт веществ и передача сигналов из внешней среды в цитоплазму. Само расположение ферментов в липидной фазе мембраны весьма существенно для их активности. Таким образом, благодаря упорядоченности и векторности ферментов и транспортных механизмов осуществляется мембранная координация множества биохимических реакций, протекающих в клетке.
Аккумуляция и трансформация энергии— одна из важнейших функций мембран. В хлоропластах зеленых растений энергия света трансформируется в энергию NADH и АТР, а в конечном счете — в стабильную энергию химических связей Сахаров, органических кислот, аминокислот. Эти ассимиляты, окисляясь затем в цитоплазме и митохондриях, освобождают энергию, необходимую клеткам для выполнения различных функций. Согласно хемиосмотической теории П. Митчелла, процесс синтеза АТР в митохондриях и хлоропластах опосредован образованием электрохимического мембранного потенциала ионов Н+ (см. рис. 1.3) за счет работы Н +-насоса.
Помимо редокс-цепей и Н+ -АТРазы в клетках имеются и другие насосы, создающие элек трохимические потенциалы: Na+ К + -АТРаза, Са2 + -АТРаза, анионная АТРаза. Как отмечает В. П. Скулачев (1978), осмотический градиент этих потенциалов — трансмембранный градиент К+ (ΔрК +) и Na+ (ΔpNa+) — нужно рассматривать как резервную форму энергии. Таким образом, в живых клетках помимо дыхательных субстратов энергия может запасаться в двух формах — в виде легко отдающих энергию химических соединений (АТР, NADH и др.) и в виде электрохимического мембранного потенциала (Δµн+, Δµк+, ΔµNa+) Все эти резервные формы энергии способны к взаимопревращениям, а также могут быть использованы на другие виды работ: химическую, осмотическую, тепловую и т. д.
Важнейшая функция мембран — рецепторно-регу-ляторная. В состав мембран входят хемо-, фото- и механорецепторы белковой природы, чувствительные к действию химических и физических факторов. Эти рецепторы воспринимают сигналы, поступающие из внешней и внутренней среды, обеспечивая адаптивные (приспособительные) ответы клеток на изменения условий существования.
Ядро. В растительной клетке ядро имеет диаметр порядка 10 мкм. Оно может быть сферическим, удлиненным или лопастным. Внутреннее содержимое ядра (нуклеоплазма) ограничено ядерной оболочкой, состоящей из двух элементарных мембран — внутренней и наружной. Ядерная оболочка пронизана порами диаметром 10 — 20 нм, через которыетранспортируются нуклеиновые кислоты и белки. С наружной мембраной ядерной оболочки могут быть непосредственно связаны элементы эндоплазматического ретикулума. В нуклео-плазме локализованы участки хроматина, состоящего из комплекса ДНК, РНК и белков. В делящихся клетках хроматин организуется в хромосомы, число которых специфично для каждого вида растений. Количество ДНК в одном ядре постоянно для каждого вида (кроме случаев мейоза и полиплоидии). Геном растений отличается от генома животных большим количеством ДНК. В нуклеоплазме ядра содержатся также ферменты и кофакторы, необходимые для обеспечения процессов репликации и транскрипции ДНК, молекулы различных РНК, ферменты, фосфорилирующие и ацетилирую-щие ядерные белки, ферменты гликолиза и др.
Ядрышко отчетливо видно в ядре под световым и электронным микроскопами. Оно формируется на определенных участках ДНК, называемых ядрышковым организатором. В хроматине ядрышка находятся участки ДНК, ответственные за синтез рибосомальных РНК (рРНК).
В целом ядро является местом хранения генетической информации клетки и репликации ДНК. В нем происходит процесс транскрипции ДНК в РНК различных типов. В тесном взаимодействии с цитоплазмой ядро участвует в обеспечении экспрессии генетической информации и контролирует процессы жизнедеятельности клетки.
Рибосомы. Рибосомы осуществляют синтез белков — трансляцию матричной, или информационной, РНК (мРНК). На электронных фотографиях они выглядят округлыми частицами диаметром 20 — 30 нм. Каждая рибосома состоит из двух нуклеопротеиновых субъединиц. В цитоплазме растительных клеток находятся 80 S рибосомы, состоящие из 40 и 60 S субъединиц, в хлоропластах — 70 S рибосомы, а в митохондриях — 78 — 80 S рибосомы, отличные от цитоплазматических и хлоропластных. Субъединицы рибосом, образованные в ядрышке, поступают в цитоплазму, где происходит сборка рибосом на молекуле мРНК.

Рибосомы в цитоплазме могут быть свободными, прикрепленными к мембранам эндоплазматического ретикулума, к наружной мембране ядерной оболочки либо образуют поли-рибосомные (полисомные) комплексы. Полирибосомы возникают потому, что одну молекулу мРНК могут одновременно транслировать несколько рибосом. Полирибосомы разрушаются при воздействии на растения неблагоприятных факторов внешней среды (например, засухи, недостатка кислорода). В процессе синтеза белка, осуществляемого рибосомами, участвуют компоненты, образованные в ядре, ядрышке, в цитоплазме, митохондриях и хлоропластах.
Пластиды. Для клеток растений типична система органоидов, называемая пластидной. Пластиды — это ограниченные двойной мембраной округлые или овальные органоиды, содержащие внутреннюю систему мембран. Пластиды могут быть бесцветными (пропластиды, лейкопласты, этиоплас ты) или окрашенными (хлоропласта, хромопласты) (см. рис. 10.4).
В меристемах присутствуют пропластиды, у которых внутренняя мембрана имеет лишь небольшие инвагинации (впячивания). Если структура пропластид сохраняется у органоидов зрелых клеток, их называют лейкопластами. В лейкопластах откладываются запасные вещества и названия они получают в зависимости от этих соединений: если запасается крахмал — амилопласты, жиры — элайопласты, белки — про-теинопласты и т. д. Этиопласты формируются при выращивании зеленых растений в темноте. При освещении они превращаются в хлоропласты, при этом внутренняя система мембран подвергается сложным перестройкам (см. рис. 10.5).
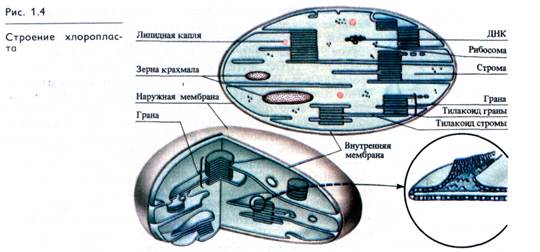
Важнейший для всего живого процесс преобразования энергии поглощенного света в химическую энергию органических веществ, синтезируемых из СО2 и Н2О (фотосинтез), осуществляется в зеленых пластидах — хлоропластах. Это овальные тела 5 — 10 мкм длиной и 2 — 3 мкм в диаметре. В одной клетке листа могут находиться 15 — 20 и более хлоропластов, а у некоторых водорослей — лишь 1 — 2 гигантских хлоропласта различной формы. Как и другие пластиды, хлоропласты обладают наружной и внутренней мембранами (рис. 1.4). Внутренняя мембрана, ограничивающая внутреннюю гомогенную среду (строму) хлоропласта, образует уплощенные инвагинации — тилакоиды, которые могут иметь форму дисков и в этом случае называются тилакоидами гран (гранальными). Несколько таких тилакоидов, лежащих друг над другом, формируют стопку — грану. Другие тилакоиды, связывающие между собой граны или не контактирующие с ними, называются тилакоидами стромы.
В мембранах тилакоидов локализованы зеленые (хлорофиллы), желтые и красные (каротиноиды) пигменты, компоненты редокс-цепей и запасания энергии, участвующие в поглощении и использовании энергии света. Биохимические системы синтеза и превращения углеводов функционируют в строме хлоропластов. В ней же может откладываться крахмал.
Хромопласты содержат в везикулах (пузырьках) стромы каротиноиды. Их присутствием объясняется окраска плодов томатов, рябины и др. В строме всех пластид обнаружены кольцевые молекулы ДНК.
При росте клетки количество хлоропластов увеличивается путем деления. При делении ламеллярная система пересекается перемычкой поперек органоида, иногда наблюдается почкование хлоропластов. Затем размер дочерних хлоропластов увеличивается. Деление хлоропластов происходит через 6 - 20 ч. и не обязательно совпадает с делением ядра. Оно может регулироваться красным светом (660 нм) и устраняется облучением дальним красным светом (730 нм). Деление останавливается также низкой температурой.
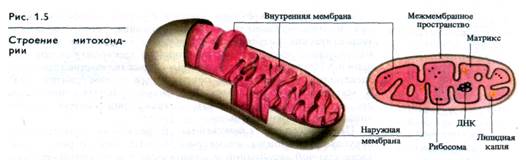
Митохондрии. В растительных клетках митохондрии представлены округлыми или гантелевидными телами диаметром 0,4 - 0,5 мкм и длиной 1-5 мкм. Количество митохондрий варьирует от десятков до 2000 на клетку, меняясь в онтогенезе клетки и в зависимости от ее функционального состояния. Органоиды ограничены наружной и внутренней мембранами толщиной 5-6 нм каждая (рис. 1.5). Внутренняя мембрана образует складки различной формы, называемые кристами (гребнями). Мембраны отделяют от цитоплазмы внутреннее содержимое митохондрий — матрикс. В матриксе содержатся рибосомы и митохондриальная ДНК, ответственные за синтез части белков митохондрий. ДНК в митохондриях имеет кольцевое строение.
Собственная генетическая система и способность мито-хондриальной ДНК к репликации позволяют митохондриям делиться независимо от деления ядра. В клетке они образуются от предсуществующих митохондрий. Продолжительность жизни (оборот) митохондрий определяется скоординированной деятельностью ядра, цитоплазмы и самих митохондрий. Полупериод жизни этих органоидов у различных растений составляет 5-10 дней, причем наружная мембрана обновляется быстрее внутренней.
В митохондриях функционируют системы аэробного дыхания и окислительного фосфорилирования, обеспечивающие энергетические потребности клеток. Во внутренней мембране митохондрий локализованы компоненты электронтранспортной цепи и АТР-синтетазные комплексы, осуществляющие транспорт электронов и протонов и сопряженный с ним синтез АТР. В матриксе располагаются ферментные системы окисления ди- и трикарбоновых кислот, а также ряд систем синтеза липидов, аминокислот и др.
Пероксисомы и глиоксисомы. В растениях присутствуют округлые органоиды диаметром 0,2—1,5 мкм, ограниченные элементарной мембраной и содержащие гранулярный матрикс умеренной электронной плотности. Они получили название микротел. В некоторых микротелах обнаруживается белковый кристаллоид, состоящий из трубочек диаметром около 6 нм. Количество микротел в клетке близко к числу митохондрий. В клетках растений обнаружены два типа микротел, выполняющих различные физиологические функции: пероксисомы и глиоксисомы.
Пероксисомы многочисленны в клетках листьев, где они тесно связаны с хлоропластами. В них окисляется синтезируемая в хлоропластах в ходе фотосинтеза гликолевая кислота и образуется аминокислота глицин, которая в митохондриях превращается в серин. В листьях высших растений пероксисомы участвуют в фотодыхании (см. рис. 3.16).
Глиоксисомы появляются при прорастании семян, в которых запасаются жиры, и содержат ферменты, необходимые для превращения жирных кислот в сахара: системы β-окисления жирных кислот и глиоксилатный цикл (см. рис. 4.11). При работе ферментных систем пероксисом и глиоксисом образуется пероксид водорода, который разрушается содержащейся в этих органоидах каталазой.
Сферосомы. Это сферические, сильно преломляющие свет образования диаметром 0,5 мкм. Они содержат липиды и поэтому их называют также липидными каплями (олеосомами). В  сферосомах обнаружены такие ферменты, как липаза и эстераза. В них хранятся запасы липидов клетки. При прорастании семян, запасающих жиры, сферосомы функционируют в комплексе с глиоксисомами в процессах глюконеогенеза.
сферосомах обнаружены такие ферменты, как липаза и эстераза. В них хранятся запасы липидов клетки. При прорастании семян, запасающих жиры, сферосомы функционируют в комплексе с глиоксисомами в процессах глюконеогенеза.
 Эндоплазматический ретикулум. Эндоплазматический ретику-лум (ЭР), или эндоплазматическая сеть (ЭС), представляет собой систему каналов, пузырьков и цистерн, ограниченную мембраной толщиной 5 — 6 нм. ЭР может содержать на своей поверхности рибосомы (гранулярный, или шероховатый, ЭР) или не содержать их (агранулярный, или гладкий, ЭР). Поверхность мембран ЭР в клетке больше других мембранных образований, а объем полостей может достигать 16% объема клетки. Эндоплазматическая сеть — очень лабильная структура. Неблагоприятные внешние воздействия (недостаток кислорода и др.) вызывают концентрические закручивания мембран ретикулума.
Эндоплазматический ретикулум. Эндоплазматический ретику-лум (ЭР), или эндоплазматическая сеть (ЭС), представляет собой систему каналов, пузырьков и цистерн, ограниченную мембраной толщиной 5 — 6 нм. ЭР может содержать на своей поверхности рибосомы (гранулярный, или шероховатый, ЭР) или не содержать их (агранулярный, или гладкий, ЭР). Поверхность мембран ЭР в клетке больше других мембранных образований, а объем полостей может достигать 16% объема клетки. Эндоплазматическая сеть — очень лабильная структура. Неблагоприятные внешние воздействия (недостаток кислорода и др.) вызывают концентрические закручивания мембран ретикулума.
 В мембранах ЭР локализованы редокс-цепи двух типов, с участием которых происходят детоксикация вредных для клетки соединений (NADPH-редуктаза и цитохром Р-450) и превращение насыщенных жирных кислот в ненасыщенные (NADH-редуктаза и цитохром Ь5). В гладком ЭР образуются углеводы, липиды, терпеноиды. В гранулярном ретикулуме синтезируются мембранные белки, ферменты, необходимые для синтеза полисахаридов клеточных стенок, структурный белок и ферменты клеточных стенок, другие секретируемые белки. По системе ЭР переносятся вещества внутри клетки. Он участвует также в межклеточных взаимодействиях у растений через плазмодесмы (см. ниже).
В мембранах ЭР локализованы редокс-цепи двух типов, с участием которых происходят детоксикация вредных для клетки соединений (NADPH-редуктаза и цитохром Р-450) и превращение насыщенных жирных кислот в ненасыщенные (NADH-редуктаза и цитохром Ь5). В гладком ЭР образуются углеводы, липиды, терпеноиды. В гранулярном ретикулуме синтезируются мембранные белки, ферменты, необходимые для синтеза полисахаридов клеточных стенок, структурный белок и ферменты клеточных стенок, другие секретируемые белки. По системе ЭР переносятся вещества внутри клетки. Он участвует также в межклеточных взаимодействиях у растений через плазмодесмы (см. ниже).
Аппарат Гольджи. В растительных клетках аппарат Гольджи (АГ) представлен диктиосомами, везикулами и межцистер-ными образованиями. Уплощенные цистерны — диктиосомы расположены пачками по несколько штук. Они ограничены мембраной толщиной 7 — 8 нм. На регенерационном полюсе АГ происходит новообразование диктиосом из мембран гладкого ЭР. На секреторном полюсе формируются секреторные пузырьки (везикулы), содержащие предназначенные для секреции вещества. В клетке растений содержатся от нескольких до сотен АГ.
В диктиосомах АГ образуются гликопротеины и гликолипиды и осуществляется накопление и мембранная «упаковка» соединений, необходимых для синтеза полимеров клеточной стенки и различных растительных слизей. С помощью везикул Гольджи углеводные компоненты доставляются к плазмалемме. Мембрана пузырьков встраивается в плазмалемму, способствуя ее росту и обновлению. Секретируемые вещества оказываются в клеточной стенке. Мембраны АГ являются связующим звеном между мембранами ЭР и плазмалеммой.
Вакуолярная система. Вакуоль — типичный органоид растительной клетки. В меристематических клетках вакуоли представлены мелкими пузырьками; для зрелых клеток характерна большая центральная вакуоль. Вакуолярная система растений формируется несколькими путями. Из расширенных цистерн ЭР образуются провакуоли, слияние которых приводит к возникновению более крупных вакуолей и созданию вакуо-лярной мембраны — тонопласта, который таким образом является производным ЭР. Тонопласт может образовывать инвагинации, что приводит к включению в вакуоль участков цитоплазмы. Гидролитические ферменты, содержащиеся в возникшей вакуоли, расщепляют полимеры до низкомолекулярных веществ.
Важную роль в становлении вакуолей играют явления автофагии. Процесс начинается с окружения мембраной ЭР участка цитоплазмы (автофагическая вакуоль). Деятельность кислых гидролаз в замкнутом мембраной пространстве приводит к деградации полимерного содержимого и поступлению воды. Вакуоли, возникшие в результате автофагии, могут сливаться с вакуолями, образовавшимися иными путями.

 Вакуолярный сок имеет сложный состав и включает органические вещества и минеральные соли. Помимо органических кислот, углеводов, аминокислот и белков, которые могут быть вторично использованы в обмене веществ, клеточный сок содержит фенолы, таннины, алкалоиды, антоцианы, которые выводятся из обмена веществ клетки в вакуоль и таким путем изолируются от цитоплазмы. Большинство ферментов вакуолей — гидролазы с оптимумом активности при кислом рН, что позволяет рассматривать вакуоли растительных клеток в качестве вторичных лизосом. Кислотность вакуолярного сока 5,0 — 6,5 единиц рН, но может быть равной 1,0 (бегония) или 2,0 (лимон).
Вакуолярный сок имеет сложный состав и включает органические вещества и минеральные соли. Помимо органических кислот, углеводов, аминокислот и белков, которые могут быть вторично использованы в обмене веществ, клеточный сок содержит фенолы, таннины, алкалоиды, антоцианы, которые выводятся из обмена веществ клетки в вакуоль и таким путем изолируются от цитоплазмы. Большинство ферментов вакуолей — гидролазы с оптимумом активности при кислом рН, что позволяет рассматривать вакуоли растительных клеток в качестве вторичных лизосом. Кислотность вакуолярного сока 5,0 — 6,5 единиц рН, но может быть равной 1,0 (бегония) или 2,0 (лимон).
Вещества доставляются в вакуоль с помощью разнообразных систем транспорта, функционирующих в тонопласте. К ним относится АТР-зависимая Н+-помпа, выносящая ионы Н+ из цитоплазмы в вакуоль (см. рис. 1.3). Ее деятельность обеспечивает поступление в вакуоль анионов органических кислот, Сахаров, а также вход и выход ионов К+. Переносчики, локализованные в тонопласте, обусловливают накопление в вакуолях аминокислот и других соединений.
Важно отметить, что вакуоль может служить местом отложения запасных белков (алейроновые зерна). Процесс вакуолизации — необходимое условие роста клеток растяжением.
Микротрубочки и микрофиламенты (цитоскелет). В наружном кортикальном слое цитоплазмы неделящихся растительных клеток локализованы микротрубочки (см. рис. 13.2). Их наружный диаметр 30 нм, внутренний — около 14 нм. Они ориентированы параллельно друг другу и перпендикулярно продольной оси клетки (см. рис. 10.6). При делении клетки микротрубочки составляют основу структуры веретена, пучки трубочек прикрепляются также к кинетохорам хромрсом. У монадных водорослей микротрубочки входят в состав жгутиков, обеспечивая их подвижность. Все микротрубочки имеют единый план строения и состоят из глобулярного кислого белка тубулина, субъединицей которого является димер, состоящий из двух глобулярных мономеров (α- и β-тубулинов). В микротрубочке димеры белка располагаются по спирали. Трубка образована 13 субъединицами тубулина. Цитоплазматические микротрубочки легко диссоциируют на субъединицы (разбираются) и собираются вновь. Для сборки микротрубочек благоприятен кислый рН среды, присутствие магния, GTP и АТР. Разборка ускоряется повышением концентрации Са+ и низкой температурой.
В цитоплазме растительных клеток обнаружены также филаментные структуры, состоящие из немышечного актина. Это сократительный белок, сходный по молекулярной массе с актином мышц и близкий ему по аминокислотному составу. Он может находиться в мономерной (глобулярный, Г-актин) или в полимерной форме двойной спирали (фибриллярный, Ф-актин). Микрофиламенты актина взаимодействуют с микротрубочками кортикального слоя и плазмалеммой. Они участвуют в пространственной организации метаболических 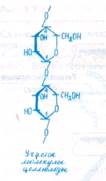
 процессов, протекающих в растворимой фазе цитоплазмы, и служат основой ее двигательной активности.
процессов, протекающих в растворимой фазе цитоплазмы, и служат основой ее двигательной активности.
Клеточная стенка. Клетки растений окружены плотной полисахаридной оболочкой, выстланной изнутри плазмалеммой. Формируется клеточная стенка на стадии телофазы во время митотического деления клеток. Клеточную стенку делящихся и растущих растяжением клеток называют первичной. После прекращения роста клетки на первичную клеточную стенку изнутри откладываются новые слои и возникает прочная вторичная клеточная стенка.
В состав клеточной стенки входят структурные компоненты (целлюлоза у растений, хитин у грибов), компоненты матрикса стенки (гемицеллюлозы, пектин, белки), инкрустирующие компоненты (лигнин, суберин) и вещества, откладывающиеся на поверхности стенки (кутин и воска). Клеточные стенки могут содержать также силикаты и карбонаты кальция.
Целлюлоза (полимер β-D-глюкозы), гемицеллюлозы (полимеры гексоз и пентоз) и пектиновые вещества (производные уроновых кислот) являются углеводными компонентами клеточных стенок. Целлюлоза и пектиновые вещества адсорбируют воду, обеспечивая оводненность клеточной стенки. Пектиновые вещества, содержащие много карбоксильных групп, связывают ионы двухвалентных металлов, которые способны обмениваться на другие катионы (Н+, К+ и т.д.). Это обусловливает катионообменную способность клеточных стенок растений. Помимо углеводных компонентов в состав матрикса клеточной стенки входит также структурный белок, называемый экстенсином. Это гликопротеин, содержащий более 20% L-оксипролина от суммы аминокислот. По этому признаку белок клеточных стенок растений сходен с межклеточным белком животных — коллагеном.
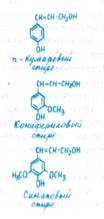
Основным инкрустирующим веществом клеточной стенки является лигнин. Интенсивная лигнификация клеточных стенок начинается после прекращения роста клетки. Лигнин представляет собой полимер с неразветвленной молекулой, состоящей из ароматических спиртов (n-кумарового, кониферилового, синапового). Разрушение и конденсация лигнина в почве— один из факторов образования гумуса.
В регуляции водного и теплового режима растений участвуют ткани, стенки клеток которых пропитаны суберином. Отложение суберина делает стенки трудно проницаемыми для воды и растворов (например, в эндодерме, перидерме).
Поверхность эпидермальных клеток растений защищена гидрофобными веществами — кутином и восками: Предшественники этих соединений секретируются из цитоплазмы на поверхность, где и происходит их полимеризация. Слой кутина обычно пронизан полисахаридными компонентами стенки (целлюлозой, пектином) и образует кутикулу. Кутикула участвует в регуляции водного режима тканей и защищает клетки от повреждений и проникновения инфекции.
В первичных клеточных стенках на долю целлюлозы приходится до 30% сухой массы стенки. Количество геми-целлюлоз и пектиновых веществ меняется в зависимости от объекта. Вместе с белками пектиновые вещества могут составлять около 30% сухой массы клетки, причем количество белка достигает 5-10%. Около 40% приходится на долю гемицеллюлоз.
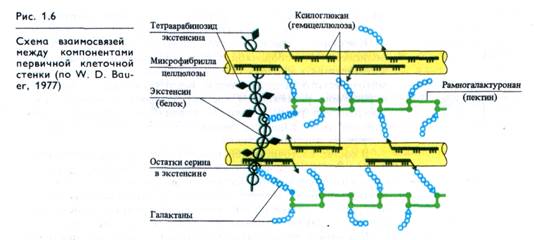
Каким образом взаимосвязаны рассмотренные компоненты в клеточной стенке? Молекулы целлюлозы взаимодействуют между собой водородными связями и собраны в микрофибриллы (рис. 1.6). Водородные связи существуют также между микрофибриллами целлюлозы и гемицеллюлозами. Большинство остальных связей в клеточной стенке ковалент-ные: между гемицеллюлозами и пектиновыми веществами, между пектиновыми веществами и экстенсином, между лигнином и целлюлозой, экстенсином и лигнином. В пектиновых полимерах, имеющих карбоксильные группы, существенную роль играют ионные связи с участием главным образом кальция. Эти взаимодействия обеспечивают прочность структуры клеточной стенки, ее эластичность и пластичность.
Благодаря контакту соседних клеток друг с другом возникает единая система клеточных стенок, получившая название апопласта. По апопласту, минуя мембранные барьеры, от клетки к клетке перемещаются вещества. Межмолекулярное пространство в фазе клеточной стенки, где осуществляются диффузия, адсорбция и освобождение водорастворимых веществ, называется кажущимся свободным пространством (см. 6.11.1).
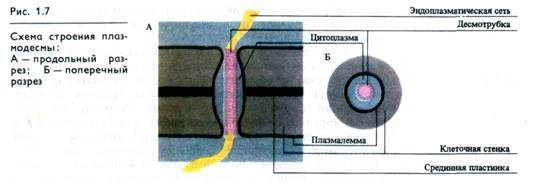 |
 Клеточные стенки растений пронизаны отверстиями — порами диаметром до 1 мкм. Через них проходят тяжи — плазмодесмы (рис. 1.7), благодаря которым осуществляются межклеточные контакты. Каждая плазмодесма представляет собой канал, выстланный плазмалеммой, непрерывно переходящей из клетки в клетку. Центральную часть поры занимает десмотрубка, состоящая из спирально расположенных белковых субъединиц. Десмотрубка сообщается с мембранами ЭР соседних клеток. Вокруг десмотрубки имеется слой цитоплазмы,которая может соединяться с цитоплазмой соседних клеток. Таким образом, связи между клетками могут осуществляться через цитоплазму, плазмалемму, ЭР и клеточные стенки. Единая система цитоплазмы клеток тканей и органов называется симпластом.
Клеточные стенки растений пронизаны отверстиями — порами диаметром до 1 мкм. Через них проходят тяжи — плазмодесмы (рис. 1.7), благодаря которым осуществляются межклеточные контакты. Каждая плазмодесма представляет собой канал, выстланный плазмалеммой, непрерывно переходящей из клетки в клетку. Центральную часть поры занимает десмотрубка, состоящая из спирально расположенных белковых субъединиц. Десмотрубка сообщается с мембранами ЭР соседних клеток. Вокруг десмотрубки имеется слой цитоплазмы,которая может соединяться с цитоплазмой соседних клеток. Таким образом, связи между клетками могут осуществляться через цитоплазму, плазмалемму, ЭР и клеточные стенки. Единая система цитоплазмы клеток тканей и органов называется симпластом.
Будучи продуктом метаболической активности протопласта, клеточная стенка выполняет функцию защиты содержимого клетки от повреждений и избыточной потери воды, поддерживает форму (за счет тургора) и определяет размер клетки, служит важным компонентом ионного обмена клетки (как ионообменник) и местом транспорта веществ из клетки в клетку внеклеточным путем (апопластный транспорт). Биогенез клеточной стенки играет важную роль в росте и дифференцировке клетки.
Тело высших растений состоит из двух главных частей — побега и корня, которые образуют главную ось растения. Побег включает стебель, листья, вегетативные почки (апикальную и боковые), цветки и плоды; корневая система — главный, боковые и придаточные корни.
Стебель выполняет опорную и проводящую функции. Он обладает также двигательной активностью (зона растяжения), часто служит местом отложения запасных веществ и в ряде случаев является органом вегетативного размножения (столоны, усы и т. д.). В связи с новыми функциями стебля, появившимися в ходе эволюции, возникли его многочисленные модификации. Например, подземные корневища, клубни, луковицы выполняют функции вегетативного размножения, хранения запасных веществ, перенесения неблагоприятных сезонных условий. Сочные фотосинтезирующие стебли суккулентов представляют собой приспособление к недостатку влаги. Функцию поддержания тела лазящих растений выполняют усики винограда и тыквенных, являющиеся модифицированными стеблями. Защитная функция свойственна колючкам стеблевого происхождения (у боярышника, гледичии).
Лист — специализированный орган воздушного питания, осуществляющий фотосинтез, газообмен и транспирацию. Видоизмененные листья могут выполнять функции запасающего органа (семядоли). У растений засушливых мест обитания листья сильно редуцируются или приобретают форму колючек (кактусы). У лазящих растений (горох, чина) листья становятся усиками, а у насекомоядных листовая пластинка превращена в ловчий аппарат.
Корень — специализированный орган почвенного питания, он поглощает воду и минеральные элементы, служит для закрепления в почве и обладает двигательной активностью (зона растяжения). Корень может также иметь запасные функции, приобретая форму корневых клубней (георгин), мясистых корней (орхидные). Выполнение новых функций приводит к возникновению корней-подпорок (баньян), ходульных корней (мангровые), дыхательных корней у болотных растений с сильным развитием аэренхимы, корней-прицепок (плющ), воздушных корней у эпифитных растений (орхидные) и других модификаций. В корне, как и в побеге, образуются специфические метаболиты, в том числе фитогормоны.
Вегетативные почки служат для нарастания побега и его ветвления.
Генеративные органы обеспечивают процесс полового размножения. Цветок представляет собой видоизмененный неразветвленный побег с ограниченным ростом, приспособленный для полового размножения с последующим образованием семян и плода. Органы цветка являются видоизмененными листьями: покровные листья формируют чашелистики и лепестки, а спорообразующие листья дают начало тычинкам и пестикам. Особенности строения цветка связаны со способами опыления. Сложная форма и яркая окраска венчика служат для перекрестного опыления насекомыми.
Каждый из перечисленных органов растительного организма построен из нескольких типов тканей, т. е. групп клеток, которые выполняют определенную физиологическую функцию и имеют сходное морфологическое строение, обеспечивающее реализацию этой функции. По функциональному значению в растениях различают следующие типы тканей: образовательные (меристемы), ассимиляционные (хлоренхима), запасающие, покровные, выделительные, механические (скелетные), проводящие и аэренхиму. Причем в каждом таком типе представлены ткани с более узкой специализацией. Например, к ассимиляционным тканям листа относятся столбчатая и губчатая паренхима, обкладка пучка. К покровным тканям — эпидермис, ризодерма, перидерма, эндодерма и др.
У растений существует несколько единых для всего организма функциональных систем, каждая из которых состоит из нескольких типов тканей и специализированных клеток. Это системы автотрофного (листья) и почвенного питания (корни), сосудистая проводящая система, которую у растений можно рассматривать как внутренний орган, опорная система (механические и другие ткани), двигательная система (зоны растяжения и участки с обратимо изменяющимся тургором клеток), половая система. Сосудистая система у растений выполняет те же функции, что и кровеносная система животных, за исключением транспорта кислорода. Диффузный характер имеют дыхательная и выделительная системы. Дыхательный газообмен облегчается с помощью межклетников, аэренхимы, устьиц и чечевичек. У большинства растений отсутствуют дифференцированные органы чувств. Нет у растений и нервной системы. Передача электрических импульсов осуществляется по проводящим пучкам.
Все эти особенности растительного организма связаны с его способом питания. Растению нет необходимости передвигаться в поисках пищи, как животным, так как СО2, вода, минеральные соли и свет есть в окружающей среде повсюду. Однако эти факторы присутствуют в «рассеянном» состоянии. Поэтому, чтобы максимально приблизиться к пище, растение должно удлинять осевые органы и развивать поверхности соприкосновения с окружающей средой. Это и определяет форму растительного организма, а также отсутствие у него специальных органов дыхания, так как растение дышит всей своей разветвленной и пластинчатой поверхностью. Медленно меняющиеся условия окружающей среды не требуют от растений быстрых двигательных реакций. Однако при необходимости в процессе эволюции у них развивается способность к быстрым движениям, как, например, у мимозы или у венериной мухоловки.
Основные функции растительного организма и их взаимосвязь можно представить в виде следующей схемы:
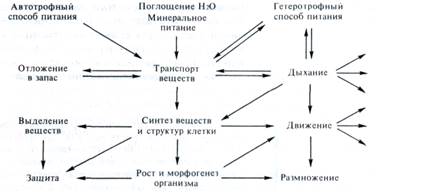
Автотрофный (фототрофый) тип питания — главная особенность растительного организма. Питание за счет фотосинтеза поддерживается корневым питанием — поглощением воды и минеральных солей. Однако все клетки и ткани растения способны питаться и гетеротрофпо. Это происходит во время прорастания (используются запасные вещества семян, клубней и т. д.) и ночью, когда фотосинтез отсутствует. К гетеротрофному питанию способны все незеленые органы. Отсюда понятно существование растений-паразитов и насекомоядных растений.
На схеме видно, что центральное место в обмене веществ целого растения занимает транспорт веществ. Этот транспорт может осуществляться по нескольким непрерывным фазам в теле растительного организма, клетки которого соединены друг с другом клеточными стенками и плазмо-десмами: по апопласту (в фазе клеточных стенок и по межклетникам), по симпласту (синцитию протопластов) и, возможно, по эндопласту (т. е. по непрерывным цистернам ЭР). Однако основным путем дальнего транспорта веществ по растению является проводящая (сосудистая) система, состоящая из ксилемы и флоэмы. Питательные вещества, поступающие по транспортным системам, пронизывающим все тело растения, при участии дыхания используются для синтеза специфических метаболитов и структур растущих и функционирующих клеток. При этом часть веществ выделяется наружу или в вакуоль. На основе деления, роста и дифференцировки клеток осуществляется морфогенез растения, а также процессы размножения. Во многих явлениях жизнедеятельности существенную роль играют двигательные реакции растений. Защитные функции растений реализуются за счет синтеза защитных веществ, часть из которых может выделяться наружу, а также благодаря формированию специальных анатомических и морфологических структур. Для всех этих процессов необходима энергия, освобождаемая при дыхании.
Таким образом, высшее растение — сложнейшая биологическая система, функциональную активность которой обеспечивают 10—15 органов, 3 — 4 десятка различных специализированных тканей, несколько десятков специализированных групп клеток. У покрытосеменных насчитывается до 80 различных типов клеток. Для сравнения отметим, что тело гидр состоит из двух тканей (эктодермы и эндодермы), в состав ее тела входят 10 различных типов клеток. У червей насчитывается до 10—12 специализированных тканей.
Строение растительного организма определяется способом его питания. Высшие растения как фототрофные организмы имеют органы воздушного (листья) и почвенного (корни) питания, а также другие физиологические системы: транспорта веществ, размножения, движения и т. д. Растительная клетка содержит все органоиды, характерные для эукариотических клеток, но в связи с фототроф-ным способом питания обладает присущей только ей пластидной системой, а также полисахаридной клеточной стенкой, двумя типами микротел (пероксисомы и глиоксисомы) и вакуолярной системой, поддерживающей тургор.
 Глава 2
Глава 2

