 2015-06-10
2015-06-10 1465
1465Гепатоцит представляет собой полярную секреторную эпителиальную клетку, имеющую базолатеральную (синусоидальную и латеральную) и апикальную (канальцевую) мембраны (см. рис. 13-5).
Образование жёлчи включает в себя захват жёлчных кислот и других органических и неорганических ионов, транспорт их через базолатеральную (синусоидальную) мембрану, цитоплазму и канальцевую мембрану. Этот процесс сопровождается осмотической фильтрацией воды, содержащейся в гепатоците и парацеллюлярном пространстве. Идентификация и характеристика транспортных белков синусоидальной и канальцевой мембран сложны. Особенно трудным является изучение секреторного аппарата канальцев, однако к настоящему времени разработана и доказала свою надёжность во многих исследованиях методика получения сдвоенных гепатоцитов в короткоживущей культуре |35|. Клонирование транспортных белков позволяет охарактеризовать функцию каждого из них в отдельности |50|.
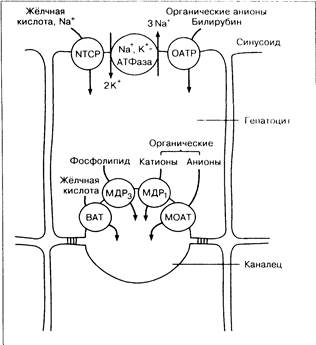
Рис. 13-5. Основные транспортные системы, участвующие в образовании жёлчи. Na+, К+-АТФаза, или натриевый насос (в центре сверху), синусоидальный котранспортный белок натрия таурохолата (NTCP — Na+-taurocholate cotransporting protein) и синусоидальный мультиспецифичный транспортный белок органических анионов (ОАТР — organic anion transporting proteon) локализуются в синусоидальной мембране. Транспортные белки канальцевой мембраны: АТФ-зависимый транспортный белок жёлчных кислот (ВАТ — ATP-dependent bile acid transporter), мультиспецифичный белок — переносчик органических анионов (МОАТ — multispecific organic anion transporter), АТФ-зависимый белок — переносчик органических катионов — белок множественной лекарственной резистентности 1 (MDR1), АТФ-зависимый транспортный белок для фосфолипидов (флиппаза) (MDR3). Другие транспортные системы: синусоидальный обменный белок для Na+/H+ и канальцевая транспортная система для бикарбонатов.
Процесс жёлчеобразования зависит от'наличия определённых белков-переносчиков в базолатеральной и канальцевой мембранах (см. рис. 13-5). Роль движущей силы секреции выполняет Na+, К +- АТФаза базолатеральной мембраны, обеспечивая химический градиент и разность потенциалов между гепатоцитом и окружающим пространством. Na+, К +- АТФаза обменивает три внутриклеточных иона натрия на два внеклеточных иона калия, поддерживая градиент концентрации натрия (высокая снаружи, низкая внутри) и калия (низкая снаружи, высокая внутри). В результате этого содержимое клетки имеет отрицательный заряд (–35 мВ) по сравнению с внеклеточным пространством, что облегчает захват положительно заряженных ионов и экскрецию отрицательно заряженных ионов. Na+, К + -АТФаза не обнаруживается в канальцевой мембране [76]. Текучесть мембран может влиять на активность фермента.
ЗАХВАТ НА ПОВЕРХНОСТИ СИНУСОИДАЛЬНОЙ МЕМБРАНЫ
Базолатеральная (синусоидальная) мембрана имеет множество транспортных систем для захвата органических анионов, субстратная специфичность которых частично совпадает (см. рис. 13-5) [58]. Характеристика белков-переносчиков ранее давалась на основании изучения клеток животных. Недавнее клонирование транспортных белков человека позволило лучше охарактеризовать их функцию [50|. Транспортный белок для органических анионов (organic anion transporting protein — OATP) является натрийнезависимым, переносит молекулы ряда соединений, включая жёлчные кислоты, бромсульфалеин и, вероятно, билирубин [49]. Полагают, что транспорт билирубина в гепатоцит осуществляют также другие переносчики [66]. Захват жёлчных кислот, конъюгированных с таурином (или глицином), осуществляется транспортным белком натрия/таурохолата (sodium/bile acid co-transporting protein — NTCP) [58].
В переносе ионов через базолатеральную мембрану участвует белок, обменивающий Na+/H+ и регулирующий рН внутри клетки. Эту функцию выполняет также котранспортный белок для Na+/HCO3–. На поверхности базолатеральной мембраны происходит также захват сульфатов, неэтерифицированных жирных кислот, органических катионов [64].
Транспорт жёлчных кислот в гепатоците осуществляется с помощью цитозольных белков, среди которых основная роль принадлежит За-гидроксистероиддегидрогеназе. Меньшее значение имеют глутатион-S-трансфераза и белки, связывающие жирные кислоты. В переносе жёлчных кислот участвуют эндоплазматический ретикулум и аппарат Гольджи. Везикулярный транспорт включается, по-видимому, только при значительном поступлении в клетку жёлчных кислот (в концентрациях, превышающих физиологические).
Транспорт белков жидкой фазы и лигандов, таких как IgA и липопротеиды низкой плотности, осуществляется посредством везикулярного трансцитоза. Время переноса от базолатеральной к канальцевой мембране составляет около 10 мин. Данный механизм ответствен только за небольшую часть суммарного тока жёлчи и зависит от состояния микротрубочек.
Канальцевая мембрана представляет собой специализированный участок плазматической мембраны гепатоцита, содержащий транспортные белки (большей частью АТФ-зависимые), ответственные за перенос молекул в жёлчь против градиента концентрации. В канальцевой мембране локализуются также такие ферменты, как ЩФ, ГГТП. Перенос глюкуронидов и глутатион-S-конъюгатов (например, билирубина диглюкуронида) осуществляется с помощью канальцевого мультиспецифичного транспортного белка для органических анионов (сапаlicular multispecific organic anion transporter — cMOAT), перенос жёлчных кислот — с помощью канальцевого транспортного белка для жёлчных кислот (canalicular bile acid transporter — сВАТ), функция которого частично управляется отрицательным внутриклеточным потенциалом. Ток жёлчи, не зависящий от жёлчных кислот, определяется, по-видимому, транспортом глутатиона, а также канальцевой секрецией бикарбоната, возможно, при участии белка, обменивающего Cl–/HCO3–.
Важная роль в транспорте веществ через канальцевую мембрану принадлежит двум ферментам семейства Р-гликопротеинов; оба фермента являются АТФ-зависимыми [63]. Белок множественной лекарственной резистентности 1 (multidrug resistance protein 1 — MDR1) переносит органические катионы, а также осуществляет выведение цитостатических препаратов из раковых клеток, обусловливая их резистентность к химиотерапии (отсюда название белка). Эндогенный субстрат MDR1 неизвестен. MDR3 переносит фосфолипиды и действует как флиппаза для фосфатидилхолина. Функция MDR3 и его важное значение для секреции фосфолипидов в жёлчь уточнены в экспериментах на мышах, у которых отсутствует mdr2-P-гликопротеин (аналог MDR3 человека) [79]. При отсутствии в жёлчи фосфолипидов жёлчные кислоты вызывают повреждение билиарного эпителия, воспаление дуктул и перидуктулярный фиброз.
Вода и неорганические ионы (в особенности натрий) экскретируются в жёлчные капилляры по осмотическому градиенту путем диффузии через отрицательно заряженные полупроницаемые плотные контакты.
Секреция жёлчи регулируется многими гормонами и вторичными мессенджерами, включая цАМФ и протеинкиназу С. Повышение концентрации внутриклеточного кальция ингибирует секрецию жёлчи. Пассаж жёлчи по канальцам происходит благодаря микрофиламентам, которые обеспечивают моторику и сокращения канальцев (рис. 13-6) [81].
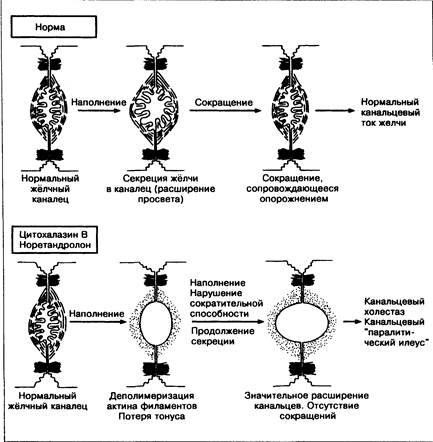
Рис. 13-6. Один из возможных механизмов развития внутрипеченочного холестаза. В норме канальцевая секреция жёлчи сопровождается сокращениями канальцев, обеспечивающими ток жёлчи. Цитохалазин и норетандролон вызывают деполимеризацию актина микрофиламентов, что ведёт к потере тонуса и сократительной способности канальцев, их расширению с последующим канальцевым «паралитическим илеусом» и развитием холестаза.
ДУКТУЛЯРНАЯ СЕКРЕЦИЯ
Эпителиальные клетки дистальных протоков вырабатывают обогащённый бикарбонатами секрет, модифицирующий состав канальцевой жёлчи (так называемый дуктулярный ток, жёлчи). В процессе секреции происходит выработка цАМФ, некоторых мембранных транспортных белков, включая белок, обменивающий Cl–/HCO3–, и регулятор трансмембранной проводимости при кистозном фиброзе — мембранный канал для Cl–, регулируемый цАМФ [16]. Дуктулярная секреция стимулируется секретином.
Предполагается, что урсодезоксихолевая кислота активно всасывается дуктулярными клетками, обменивается на бикарбонаты, рециркулирует в печени и в последующем вновь экскретируется в жёлчь («холегепатический шунт»). Возможно, этим объясняется холеретическое действие урсодезоксихолевой кислоты, сопровождающееся высокой билиарной секрецией бикарбонатов при экспериментальном циррозе [25].
Давление в жёлчных протоках, при котором происходит секреция жёлчи, в норме составляет 15— 25 см вод. ст. Повышение давления до 35 см вод. ст. приводит к подавлению секреции жёлчи, развитию желтухи. Секреция билирубина и жёлчных кислот может полностью прекращаться, при этом жёлчь становится бесцветной {белая жёлчь) и напоминает слизистую жидкость.
Синдром холестаза








