 2015-06-14
2015-06-14 760
760Одной из таких методик является электроэнцефалографическая, получившая за последние 10 – 20 лет весьма широкое распространение и обладающая целым рядом достоинств в качестве метода исследования высшей нервной деятельности человека. В принципе возможны два пути использования электроэнцефалографических показателей при изучении условных рефлексов.
Один из них заключается в выработке таких условных рефлексов, в которых регистрируется реакция какого-либо внешнего, периферического эффектора и одновременно прослеживаются частотно-амплитудные изменения электроэнцефалограммы в интересующих исследователя отведениях.
Другой путь предусматривает использование в качестве эффектора самой корковой ритмики: применяя в качестве подкрепления оптический раздражитель, а в качестве условного стимула, как обычно, любое сенсорное воздействие (например, звук), можно вызвать реакцию десинхронизации корковой ритмики – блокаду, депрессию или подавление альфа-ритма – в ответ на это сенсорное воздействие, хотя до сочетаний оно не имело этой способности. Указанное явление обладает всеми чертами выработанной реакции, свойственными классическому условному рефлексу, кроме, пожалуй, одного (не считая необычного эффекторного выражения); но об этом речь будет идти ниже. Предметом наших исследований служил именно этот последний вид условнорефлекторных изменений электроэнцефалограммы (ЭЭГ).
Первые заключения о возможности электрокоркового «обусловливания» этого типа были сделаны Г. Дюрупом и А. Фессаром (G. Dump, A. Fessard, 1935) и А. Лумисом и др. (A.L. Loomis et al., 1936) почти три десятилетия тому назад. Вслед за ними Л. Трэвис и Дж. Игэн (L. Е. Travis, J.P. Egan, 1938), X. Джаспер и К. Шагасс (Н. Jasper, С. Shagass, 1941), Дж. Нотт и К – Генри (J.R. Knott, С.Е. Henry, 1941) и целый ряд других авторов начали экспериментальное исследование проблемы электрокортикальных условных связей. Было показано, что образованные путем сочетания звука со светом условные реакции обладают генерализованностью, могут быть угашены и вновь восстановлены, что к ним может быть выработана дифференцировка, а сами они могут быть сделаны запаздывающими или следовыми и т. д. (К. Motokawa, 194t); К. Iwama, 1950; К. Motokawa, В. Huzimori, 1949; К. Iwama, М. Abe, 1952; F. Morrell, Н. Jasper, 1956; В.А. Кожевников, А.М. Марусева, 1949; А. Гасто и др., 1957; Н. Иошии и др., 1962).
Новые идеи относительно внутреннего механизма этих реакций возникли вслед за открытием Г. Моруцци и X. Мэгуном (G. Moruzzi, Н. W. Magoun, 1949) восходящего активирующего влияния мезенцефалической ретикулярной формации. Было установлено, что десинхронизация корковой ритмики является специальным результатом проведения возбуждения по экстралемнисковым путям; если импульсы, идущие от сенсорных систем по классическим путям, приносят в кору специфическую сенсорную информацию, то по коллатералям через ретикулярную систему ствола в кору поступает «неспецифическое» (в отношении сенсорной модальности – П.К. Анохин, 1962) возбуждение, роль которого заключается в диффузной активации и повышении тонуса коры, что электрографически и выражается в десинхронизации корковой ритмики (реакция пробуждения, или активации).
Д. Росси, А. Цанкетти (1960), суммируя экспериментальные данные, показали, что именно ретикулярная система генерирует те импульсы, которые вызывают корковую активацию: при перерезке ретикулярных путей поток возбуждения, идущий через лемнисковые проекции, уже не приводит к блокаде доминирующего в ЭЭГ ритма; «обрезкой» участка коры и перерезкой мозолистого тела реакция активации не уничтожается; в то же время у препарата «изолированный мозг» активация не может быть вызвана с тех зон коры, стимуляцией которых она обычно запускается.
Можно, следовательно, считать твердо установленным, что корковая активация, проявляющаяся в десинхронизации регистрируемой ритмики, однозначно связана с деятельностью ретикулярной формации ствола. В таком случае условная десинхронизация, вызываемая сочетаниями нейтрального стимула с безусловно оптическим раздражителем, представляет собой, очевидно, эффект условнорефлекторно вызванного возбуждения ретикулярной системы, и в этом явлении исследователь, по существу, сталкивается с временной связью, одним из членов которой являются структуры ретикулярной формации.
Означает ли это, что замыкание условной связи типа «звук плюс свет» и происходит на подкорковом уровне, на уровне нейронов медиальной части ствола, как это полагают, скажем, Н. Иошии, X. Пруво и X. Гасто (N. Ioshii et al., 1957)? По-видимому, такая возможность не исключена; по крайней мере, фактам, представленным указанными авторами, до сих пор не дано экспериментального опровержения. Подчеркнем, что речь идет о реакциях, вырабатываемых на зрительном подкреплении, реакциях типа «звук плюс свет», являющихся, по выражению П.К. Анохина, лишь «моделью» условного рефлекса, лишенной «самого решающего фактора истинного условного рефлекса – физиологической активации от врожденной деятельности, всегда аффективно окрашенной и захватывающей глубокие подкорковые образования» (1958, с. 16).
Трудно сомневаться в том, что «истинные», биологически значимые условные рефлексы действительно, как о том свидетельствуют многочисленнейшие факты, осуществляются при обязательном участии корковых структур. Но почему следует непременно предполагать ведущую роль коры в реализации условных реакций совсем другого, по-видимому низшего, типа реакций, вполне искусственных и совсем не принимающих участия в установлении «деловых» (по выражению И.П. Павлова) отношений со средой?
Каков бы ни был внутренний морфологический механизм замыкания связей, о которых идет речь, важно еще раз заметить, что в эти связи, по-видимому, вовлекаются в качестве «эффектора» ретикулярные образования ствола и, таким образом, в ЭЭГ реакциях типа «звук плюс свет» мы имеем дело с «обусловливанием» функций восходящей активирующей системы.
Обратимся теперь к конкретному экспериментальному материалу, полученному нами в ходе исследования индивидуальных вариаций выработки условных электрокорковых рефлексов.
Эксперименты ставились в следующих условиях. Испытуемый находился в звукоизолированной, экранированной и затемненной камере, в положении лежа, с открытыми глазами. Применялось биполярное затылочно-височное или затылочно-теменное отведение. Электродами служили серебряные диски диаметром 1 см, обернутые ватой, смоченной в физиологическом растворе. Регистрация биотоков производилась с помощью 4‑канального электроэнцефалографа.
Использовались два типа раздражений: звуковые и световые – первые как условный, вторые как подкрепляющий стимул. Звуковым раздражителем служил в одной серии опытов прерывистый звук 500 Гц (частотой 10 посылок в 1 с) громкостью около 70 дБ от среднего порога, подаваемый через динамик от звукового блока фото-фоностимулятора ФФС 01; в другой серии применялся непрерывный звук той же высоты и громкости, подаваемый от звукового генератора.
Подкреплением в обеих сериях являлся свет яркостью около 40 лк на белое, падающий через окно в стенке камеры на экран, расположенный перед глазами испытуемого. Длительность звукового сигнала составляла 4 с; при сочетаниях подкрепление присоединялось через 1 с (в первой серии) или через 2 с (во второй серии) и прекращалось вместе с условным раздражителем; интервалы обеспечивались системой реле времени, запускавшейся экспериментатором. Для выявления условной реакции в некоторых предъявлениях подкрепление опускалось.
Схема сочетаний и изолированных предъявлений условного стимула была строго фиксированной: пробы условной реакции делались после 2, 5, 7, 11, 14, 18, 22 и 25‑го сочетаний звука со светом. Всего, таким образом, производилось 8 предъявлений изолированного условного раздражителя (у некоторых испытуемых первой серии это число было сокращено до 7). Интервалы между сочетаниями и пробами составляли около 20 – 30 с.
Учет реакций производился следующим образом. Для каждого изолированного предъявления условного сигнала измерялась длительность блокады альфа-ритма в секундах от начала блокады (во время действия раздражителя) до восстановления альфа-ритма, независимо от того, где это восстановление происходило: еще до окончания действия раздражителя или после. Критерием восстановления альфа-ритма служило появление группы альфа-волн в течение 1 с и более.
Испытуемыми служили лица обоего пола в возрасте от 18 до 30 лет, в основном студенты вузов. В экспериментах по выработке условных реакций участвовали только те испытуемые, у которых альфа-ритм был достаточно выраженным, т. е. примерно 75–80 % первоначального контингента.
Всего были выполнены две серии опытов по электрокорковым условным реакциям. Хотя в некоторых методических деталях, указанных выше, вторая серия несколько отличалась от первой, это отличие, судя по результатам выработки реакций, не было существенным; по крайней мере, статистический анализ не вскрыл значимых различий между средними количествами условнорефлекторного эффекта, подсчитанными раздельно для первой и второй серий. Это дает нам право объединить результаты обеих серий и рассматривать полученные в них результаты в целом.
Какой же характер носят эти результаты?
На рис. 4 сплошной линией представлены усредненные для двух групп (39 испытуемых) данные об условнорефлекторном эффекте блокады альфа-ритма как функции числа сочетаний (точнее, числа изолированных предъявлений условного сигнала). Можно видеть, что если при первой пробе условного сигнала длительность реакции составляет примерно 5 с, то при последней она не достигает и 2 с; это значит, что при первой пробе в среднем наблюдается как наличное, так и следовое блокирование альфа-ритма, в то время как в последней пробе – и значительно раньше – условный сигнал вызывает подавление альфа-ритма только в пределах своего звучания. В целом кривая имеет тенденцию к монотонному приближению к оси абсцисс, прерываемому только некоторым «всплеском» на седьмой пробе – внезапным увеличением условной реакции на этом предъявлении, которое имело место у 12 испытуемых как из первой, так и из второй серии. Таким образом, кривая «выработки» условной десинхронизации напоминает, скорее, кривую угашения: условно-рефлекторный эффект максимален на первом предъявлении условного сигнала и минимален в конце серии сочетаний звука со светом.
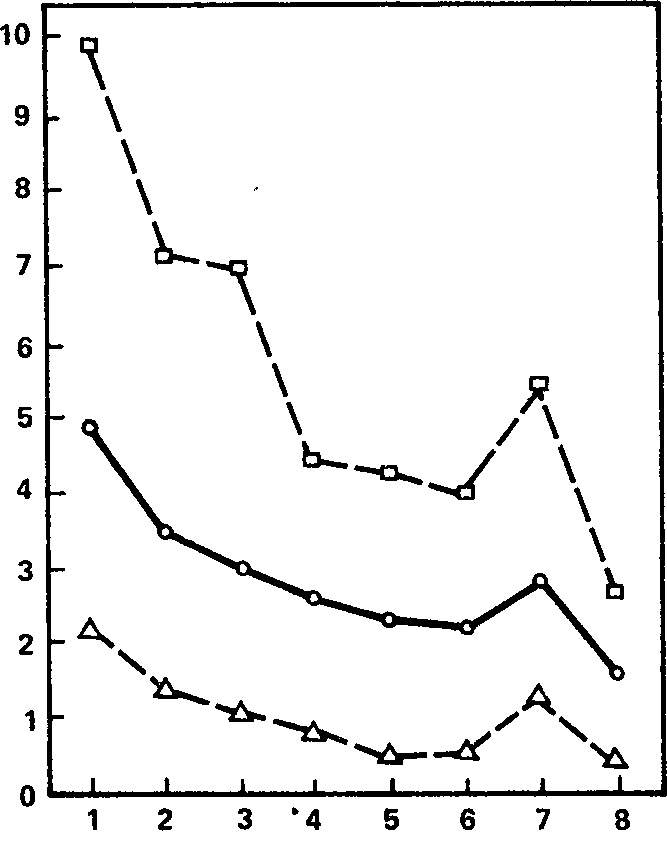
Рис. 4. Динамика условного эффекта при выработке условных электрокорковых реакций типа «звук плюс свет». Сплошная линия – средние данные для всей выборки, штриховая линия с квадратами – средние данные для группы лиц с лучшими показателями условной десинхронизации, штриховая линия с треугольниками – средние данные для группы с худшими показателями. Ось абсцисс – порядковый номер предъявления условного раздражителя; ось ординат – длительность условной реакции десинхронизации (в с).
Может быть, это уменьшение связано с тем, что длительность условной реакции измерялась при изолированных, неподкрепляемых предъявлениях условного раздражителя? Эти пробы делались сравнительно часто, что, конечно, не могло не способствовать угашению условнорефлекторного ответа. Однако данные тех авторов, которые применяли другие способы подсчета условнорефлекторного эффекта, не связанные с пропуском подкрепления, заставляют отказаться от этого предположения.
Так, К. Уэллс и X. Вулф (С.Е. Wells, Н. G. Wolff, 1960), регистрируя в качестве условной реакции блокаду альфа-ритма в интервале между началом условного и началом подкрепляющего раздражителей и строя график частоты проявления этой реакции как функции количества сочетаний, нашли постепенное снижение числа условных реакций к концу опыта (правда, в последних сочетаниях наблюдалось некоторое повышение числа реакций). Аналогичные данные приводят Дж. Стерн и др. (J.A. Stern et al., 1961 а), а также С. Виссер (S.L. Visser, 1961, 1963), использовавшие тот же метод учета условного эффекта. Обсуждая свои данные, Дж. Стерн и др. выдвигают предположение о том, что уменьшение условного эффекта в ходе сочетаний представляет собой результат развития адаптации к комплексному раздражителю, а С. Виссер, вслед за Г. Уолтером, предлагает отличать эти реакции от условных, обозначив их термином «контингентные», или «сочетанные» (contingent).
По существу, и то и другое, очевидно, не лишено оснований, но только нужно иметь в виду, что в школе И.П. Павлова давно уже возникло название для реакций подобного типа: в том случае, когда в качестве подкрепления использовался раздражитель, привлекающий внимание испытуемого (вызывающий ориентировочный рефлекс), условная реакция, вырабатывавшаяся на его основе, обозначалась как условно-ориентировочная.
Есть много оснований думать, что и ЭЭГ реакции типа «звук плюс свет» относятся к тому же классу условно-ориентировочных реакций. Действительно, блокада альфа-ритма большинством авторов, начиная с Бергера, рассматривается как биоэлектрическое выражение концентрации внимания. Так, X. Джаспер говорит: «…изменения альфа-ритма вызываются не самим сенсорным раздражителем, а вниманием к нему… Подавление альфа-ритма при ожидании подкрепления тоже связано не с афферентным притоком как таковым, а с состоянием внимания» (Н.Н. Jasper, 1949, с. 418; см. также: Д.Б. Линдслей, 1960). Не вызывает также сомнения, что с нейрофизиологической стороны реакция угнетения альфа-ритма есть не что иное, как один из компонентов ориентировочной реакции организма (Г.В. Гершуни, 1949; Е.Н. Соколов, 1958 б). Исходя из этого, можно ожидать, что снижение эффективности подкрепляющего зрительного раздражителя приведет и к снижению уровня вырабатываемой на его основе условной реакции и что, напротив, повышение эффективности подкрепления как стимула, вызывающего и поддерживающего ориентировочную реакцию, окажет положительное влияние на величину условного ответа.
Оба эти предположения подтверждаются в опыте. Так, измерив длительность блокады альфа-ритма в ответ на каждое сочетание звукового и светового раздражителей и вычислив соответствующие средние, можно построить график изменений этого показателя как функции числа комбинированных предъявлений. Динамика этих изменений показана на рис. 5, где сплошная линия соединяет соответствующие ординаты, вычисленные для 20 испытуемых второй серии. Здесь же штриховой линией изображена динамика изменений условной реакции, выявляемой при изолированных пробах звукового раздражителя.
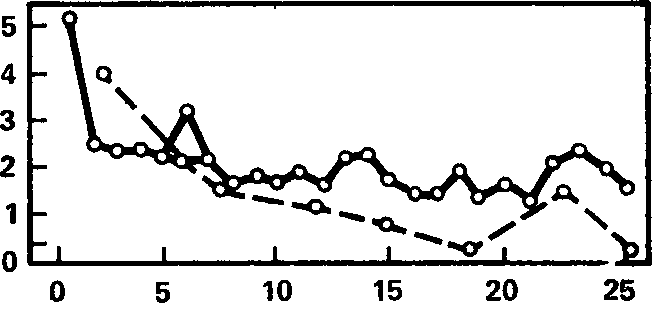
Рис. 5. Сопоставление динамики реакции десинхронизации при сочетаниях звука со светом (сплошная линия) с динамикой этой реакции при изолированных предъявлениях условного звукового раздражителя (штриховая линия). Ось абсцисс – порядковый номер сочетаний; ось ординат – длительность реакции десинхронизации (в с).
На рис. 5 видно, что десинхронизация, вызываемая световым раздражителем, резко уменьшается после первого же сочетания и в дальнейшем имеет тенденцию к постепенному сокращению[27], прерываемому, однако, кратковременными подъемами.
Сравнивая эту кривую с кривой условной реакции, можно заметить определенный параллелизм в динамике их развития: вторая кривая, так же как и первая, обнаруживает значительное падение после первой пробы, затем идет постепенное снижение, а в конце наблюдается подъем условного эффекта, явно связанный с подъемом кривой реакции на подкрепление. Подобный же параллелизм между реакциями на звук и свет отмечают уже упоминавшиеся Дж. Стерн и др. (J.A. Stern et al., 1961 а). Таким образом, величина условной десинхронизации на звук оказывается в зависимости от «безусловной» десинхронизации на свет, являющийся подкрепляющим раздражителем.
Эта же зависимость выявляется и при подсчете корреляции между средней длительностью условной блокады альфа-ритма для данного испытуемого и средней длительностью свойственной ему реакции на свет, имеющей место при сочетаниях. В первой серии коэффициент корреляции рангов между этими показателями был 0,79 (р < 0,001), во второй – коэффициент корреляции, вычисленный по методу моментов, составлял 0,509 (р <0,05). Как видим, зависимость между величиной реакции десинхронизации и эффективностью подкрепления подтверждается и статистическими показателями связи.
Наконец, совершенно отчетливо роль «ориентировочной» эффективности подкрепляющего стимула при выработке реакций типа «звук плюс свет» раскрывается в тех формах опытов, когда особыми приемами внимание испытуемых с помощью инструкции экспериментатора сосредоточивается на подкрепляющем зрительном воздействии, т. е. когда этому воздействию придается сигнальное значение (П.К. Анохин, 1958; С.А. Каразина, 1958). В наших опытах (В.Д. Небылицын, 1961) в качестве такого активирующего подкрепления мы использовали предъявление на экран перед испытуемым различных сюжетных изображений, предварительно предлагая испытуемому запомнить детали этих изображений. Было найдено, что применение активирующего подкрепления у подавляющего большинства испытуемых способствует стойкому поддержанию условной реакции на уровне, достаточном для проведения задуманных испытаний и процедур. Подробнее техника и результаты использования активирующего подкрепления изложены ниже, в гл. VII.
Итак, судя по средним данным, полученным для значительной выборки испытуемых, условная десинхронизация обнаруживает явную тенденцию к падению уже в ходе сочетаний, и это падение, как можно думать, связано с недостаточностью сигнального значения светового раздражителя, обычно применяемого в качестве ориентировочного подкрепления.
Существуют ли здесь индивидуальные различия, и если да, то какой характер они носят? На этот вопрос отвечает рис. 4, на котором, наряду с общим графиком динамики условных реакций, нанесены (штриховой линией) две кривые, характеризующие: одна – динамику условного эффекта для 10 испытуемых с лучшими средними показателями, а другая-то же для 10 испытуемых с худшими средними показателями условной реакции. Видно, что ординаты первой кривой примерно в 5 – 6 раз выше, чем ординаты второй кривой. Эти данные в определенной степени отражают диапазон индивидуальных различий в функции замыкания электрокорковых временных связей.
На рисунке видно также, что как первая, так и вторая кривая демонстрирует в общем тот же характер динамики условнорефлекторного показателя, что и средняя кривая. Следовательно, уменьшение условного эффекта к концу серии сочетаний не связано с влиянием результатов, показываемых какой-либо одной группой испытуемых, а представляет собой общую закономерность, одинаково имеющую место у лиц как с высокими, так и с низкими значениями условного эффекта.
Несколько индивидуальных кривых приведено на рис. 6. Эти кривые обнаруживают некоторые особенности, сглаживаемые при усреднении. Так, для испытуемых с высокими показателями условной реакции характерны кривые с большими колебаниями эффекта от пробы к пробе, так что иногда реакция заканчивается еще до окончания действия условного сигнала, а в других пробах она измеряется десятками секунд. Кроме того, у некоторых испытуемых условный эффект вообще не выявляется или возникает лишь в отдельных предъявлениях условного раздражителя. Есть и такие испытуемые, у которых условная реакция сохраняет примерно одну и ту же величину во всех измерениях.
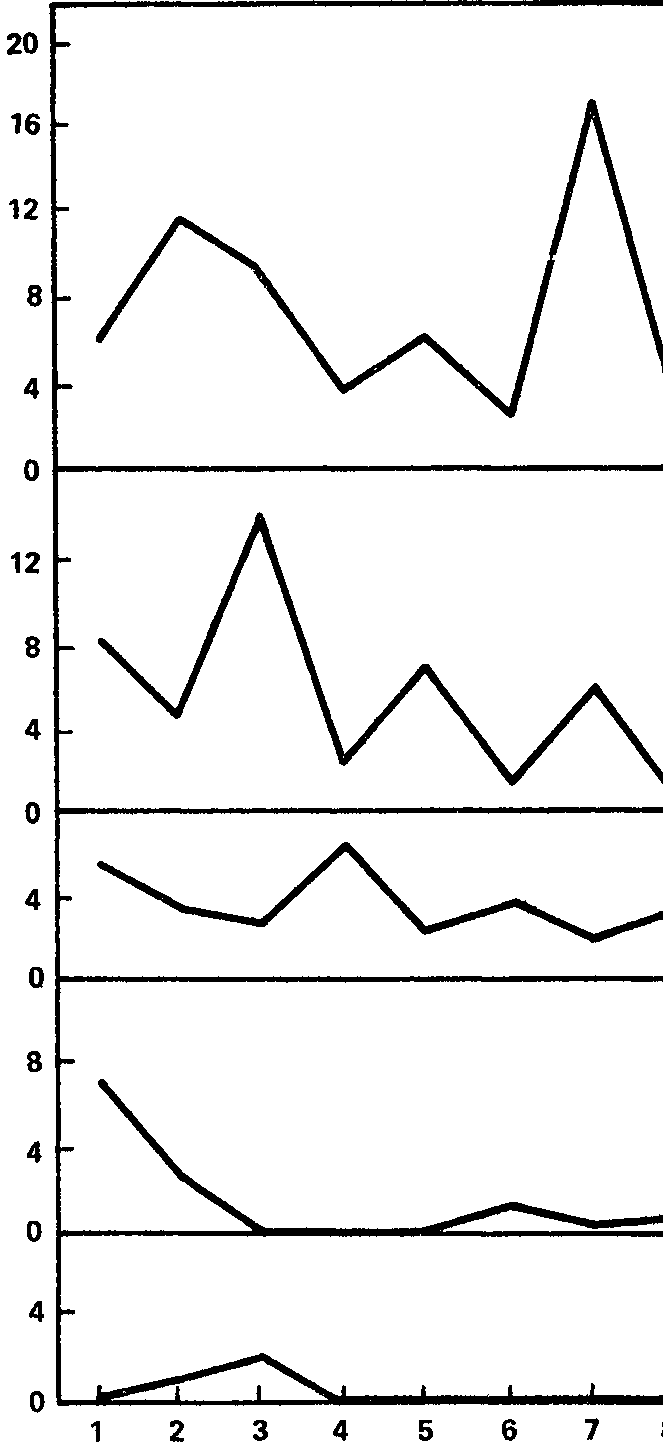
Рис. 6. Индивидуальные графики выработки условных электрокорковых реакций типа «звук плюс свет». Ось абсцисс – порядковый номер предъявлений условного сигнала; ось ординат – длительность реакции блокады альфа-ритма (в с)
В соответствии с общим представлением о структурных механизмах связей такого типа индивидуальные различия в их замыкании, по-видимому, должны пониматься как индивидуальные вариации в той легкости, с которой вступают во временную связь активирующие ретикулярные образования, ответственные за феномен десинхронизации. Диапазон этих различий весьма обширен: на одном полюсе находятся лица, у которых нет даже и признака выработки условной десинхронизации, на другом – такие испытуемые, у которых этот эффект возникает немедленно и поддерживается до самого конца серии сочетаний.








