 2015-06-14
2015-06-14 1784
1784Одним из наиболее существенных – но в то же время и неразработанных – аспектов теории свойств нервной системы является проблема соответствия между их показателями в том случае, когда последние так или иначе относятся к различным отделам (зонам, центрам, участкам) центральной нервной системы. Вопрос этот приобрел значительную актуальность с тех пор, как разными авторами были обнаружены явные расхождения в типологических оценках одного и того же индивида в зависимости от особенностей применяемой методики и, в частности, от модальности используемой в опыте стимуляции. Эти факты не могли не привлечь внимания, так как проблема разного выражения свойств нервной системы в разных структурах одного и того же мозга имеет далеко идущие теоретические последствия как для физиологии высшей нервной деятельности, так и для психологии.
Попытка классифицировать возможные причины расхождений в диагнозе того или иного свойства нервной системы приводит к выделению двух основных классов явлений, которые могут быть ответственными за эти расхождения, двух основных звеньев рефлекторной деятельности, вариации каждого из которых могут теоретически повлиять на оценку свойства нервной системы. Речь идет об афферентной и эфферентной сторонах рефлекторной деятельности, о зависимости диагноза свойств нервной системы, с одной стороны, от модальности применяемой стимуляции, а стало быть, от параметров афферентных структур мозга, а с другой стороны, от эффекторно-го выражения регистрируемой реакции и, значит, от особенностей, присущих эфферентным мозговым центрам.
Поскольку, однако, аффрентная сторона каждого условно-рефлекторного акта содержит два компонента – условный сигнал и подкрепление, возникает дополнительная возможность таких расхождений в диагнозе свойств нервной системы, которые обусловлены качествами. каждого из этих двух компонентов в отдельности, точнее, вариациями параметров тех нервных структур, к которым адресуются эти компоненты условнорефлекторного акта. Следовательно, открывается возможность таких несовпадений в оценке того или иного свойства, которые определяются либо различиями в сенсорной модальности условного раздражителя, либо различиями в биологической модальности раздражителя, применяемого в качестве подкрепления. В первом из этих случаев дело сводится, очевидно, к различиям характеристик данного свойства от одного анализатора к другому, а во втором – к подобным же различиям между вовлекаемыми в деятельность, – по-видимому, подкорковыми – центрами безусловных реакций.
Таким образом, «парциальность» проявлений свойств нервной системы, в сущности, возможна в трех основных аспектах деятельности нервной системы: 1) при регистрации разных эффекторных выражений рефлекторной деятельности (эффекторный аспект), 2) при применении условных раздражителей различной сенсорной модальности (условно-афферентный, анализаторный аспект) и 3) при использовании различных подкрепляющих воздействий (безусловно-афферентный, подкорковый аспект). По каждому из этих аспектов в литературе имеется определенный материал, к сожалению, не всегда достаточно обширный. Большей частью он свидетельствует о том, что парциальность того или иного рода в проявлениях свойств нервной системы существует не только теоретически, но и фактически. Однако это относится в основном к афферентным аспектам проблемы парциальности. Что же касается возможных проявлений парциальных особенностей в зависимости от эффекторной функции, то здесь материал слишком невелик, чтобы на этот вопрос можно было дать сколько-нибудь определенный ответ.
Вопрос о зависимости типологического диагноза от характера эффекторного выражения реакции был исследован в небольшой, проведенной всего на двух собаках работе М.С. Алексеевой (1953 б), в которой испытание по малому стандарту проводилось сначала по секреторной (слюнной) методике, а затем по двигательной. В обоих случаях условные раздражители в стереотипе относились к одним и тем же анализаторам (слуховому и зрительному), а безусловным раздражителем было подкармливание мясо-сухарным порошком. Таким образом, условия опыта во всем, исключая регистрируемую функцию, были приблизительно одинаковы. Практически одинаковыми оказались и результаты определения свойств нервной системы на основе секреторной и двигательной функций (у одной собаки – в большей, у другой – в меньшей степени).
Отсюда можно было бы сделать вывод о том, что эфферентный фактор не влияет на определение свойств нервной системы и что данная комбинация условного и подкрепляющего раздражений дает одни и те же результаты в опыте по определению этих свойств независимо от той функции, которая при этом регистрируется. Однако такой вывод, который был бы очень заманчивым, так как позволил бы пренебречь ролью эфферентных образований в вариациях свойств нервной системы, оказался бы, очевидно, преждевременным, так как совпадение результатов, полученное только на двух животных, не может вести к заключениям общего порядка.
Между тем не существует принципиальных препятствий к тому, чтобы подобное сопоставление было проведено в более широких масштабах, с использованием статистических индикаторов связи. Так, на человеке можно было бы сравнить при одних и тех же условном и подкрепляющем раздражителях (звук или какой-нибудь другой стимул плюс свет) показатели свойств нервной системы, получаемые путем регистрации таких различных в эффекторном выражении функций, как подавление альфа-ритма ЭЭГ, снижение чувствительности зрительного анализатора, сужение зрачка и, возможно, мигание. К сожалению, подобные работы до сих пор не были организованы ни на человеке, ни на животных. Поэтому вопрос о зависимости диагноза свойств нервной системы от эффекторного выражения регистрируемой функции, от эфферентного фактора следует считать открытым.
Гораздо больше внимания исследователей привлекал безусловно-афферентный аспект проблемы парциальных особенностей свойств нервной системы. Вопрос о роли подкрепления как возможной причины этих особенностей был подвергнут изучению в ряде работ сотрудников А.Г. Иванова-Смоленского, которые сопоставляли результаты определения свойств нервной системы у детей при ориентировочном, пищевом и оборонительном подкреплениях. Собственно, как уже отмечалось, с нашей точки зрения, эти авторы исследовали только один параметр деятельности нервной системы – динамичность нервных процессов, поэтому речь идет о роли безусловно-афферентного фактора в проявлениях динамичности возбуждения и торможения. Как правило, условными раздражителями служили стимулы одной и той же звуковой модальности, а эффекторным выражением условного ответа была двигательная реакция.
Материал, полученный несколькими авторами, видимо, довольно ясно свидетельствовал о существовании значительных расхождений в динамике рефлекторной деятельности при использовании подкреплений различного типа (З.Л. Синкевич, 1930; И.И. Короткий, 1930; О. Капустник, В. Фадеева, 1930; Л.Е. Хозак, 1933). Так, в работе Л.Е. Хозак (1933) было найдено, что полное совпадение индикаторов по всем трем видам подкрепления имело место только в 22 % случаев, совпадение по двум видам – в 50 % случаев. Следует, правда, иметь в виду, что автором были подвергнуты сопоставлению целостные характеристики «типов замыкательной деятельности», т. е. соотношений индикаторов образования положительных и тормозных связей, иными словами, характеристики баланса возбудительного и тормозного процессов по динамичности. Если же по данным работы Л.Е. Хозак провести раздельное сопоставление индикаторов динамичности возбуждения и индикаторов динамичности торможения (что будет правильнее, так как эти два вида динамичности, видимо, представляют собой два различных фактора деятельности нервной системы – см. об этом предыдущую главу), то оказывается следующее: показатели выработки положительных связей совпадают по трем видам подкрепления в 73 % всех случаев, а по двум – в 75 %, показатели же выработки дифференцировки при сопоставлении трех видов подкрепления совпадают в 55 % случаев, а при сопоставлении двух – в 70 % всех случаев. Как видим, процент совпадений достаточно велик, но в то же время и расхождения наблюдаются довольно часто – примерно в одной трети случаев. Это позволяет сделать вывод, согласно которому характеристика динамичности каждого из нервных процессов в отдельности имеет тенденцию в большинстве случаев оставаться постоянной независимо от рода реакции, используемой в качестве подкрепления, но у многих индивидов при разных подкреплениях она все же оказывается различной.
Интересно, что М.С. Алексеева (1953 а), сопоставившая, – к сожалению, только на трех собаках – результаты испытаний свойств нервной системы при пищевом и кислотно-оборонительном подкреплениях, приходит к выводу о том, что условные связи при этих подкреплениях образуются в общем с одинаковой скоростью. Таким образом, и в опытах на животных проявления динамичности нервных процессов имеют, видимо, тенденцию к совпадению даже при столь различных видах подкрепления, как пищевое и оборонительное (эффектор – слюнная железа – был одним и тем же в обеих формах опыта, условные сигналы относились к одним и тем же анализаторам).
Что касается других свойств нервной системы, то данные автора позволяют, видимо, предполагать, что случаи расхождения в их диагнозе на базе различных подкреплений, вообще говоря, возможны; так, у одной из собак противоположный эффект по сравнению с другой дало применение кофеиновой пробы для определения силы нервной системы, а у другой оказались отличными результаты испытания подвижности при помощи переделки.
Подытоживая имеющиеся в литературе данные относительно роли безусловно-афферентного фактора в диагнозе свойств нервной системы, можно заключить, что случаи расхождений в оценке тех или иных свойств нервной системы в зависимости от рода подкрепления, вероятно, могут иметь место, будучи, по предположениям упомянутых авторов, обусловлены интраиндивидуальными вариациями в возбудимости соответствующих безусловных центров. Насколько верно это последнее предположение, судить трудно, так как непосредственные свидетельства в его пользу отсутствуют, однако допустить в принципе внутримозговые вариации возбудимости (или каких-либо иных качеств, ответственных за параметры рефлекторной функции) нервных структур, служащих основой для функционирвания условных реакций, очевидно, возможно. В этом случае в классификацию свойств нервной системы пришлось бы ввести в качестве «поправочного» (а может быть, и самостоятельного) фактора функцию не только корковых, но и субкортикальных образований, и не только неспецифических, играющих, как видно, существенную роль в общей характеристике баланса по динамичности (В.Д. Небылицын, 1964 б), но и тех из них, которые ответственны за специфические формы безусловнорефлекторного реагирования.
Перейдем теперь к рассмотрению того аспекта интраиндивиду-альных вариаций свойств нервной системы, который был ранее обозначен как условно-афферентный, или анализаторный, и который, пожалуй, наиболее «прозрачен» с точки зрения возможностей его нейрофизиологического толкования. Этот условно-афферентный аспект раскрывается в том случае, когда при постоянстве двух компонентов условной реакции – подкрепления и реакции – вариации модальности третьего компонента, именно условного раздражителя, приводят к различному диагнозу тех или иных свойств нервной системы. Здесь наиболее вероятно предположение о том, что причиной расхождений в диагнозе являются различия функции анализаторов в их высших корковых структурах.
О том, что такие различия в принципе возможны, а у животных в связи с видовой спецификой условий обитания даже закономерны, говорят данные Э.Г. Вацуро и его сотрудников, которые обнаружили различные уровни ряда параметров нервных процессов в разных анализаторах у обезьян и собак. Так, было найдено, что у шимпанзе условные связи в несколько раз быстрее образуются в том случае, когда условные стимулы адресуются к кинестетическому анализатору по сравнению со зрительным (Э.Г. Вацуро, 1945). Если условные раздражители имели одинаковую физиологическую эффективность, этот факт может означать более высокую динамичность возбудительного процесса в кинестетическом анализаторе у шимпанзе.
В исследовании, проведенном на собаках, подвижность процессов, протекающих в слуховом анализаторе этих животных, была найдена более высокой по сравнению с подвижностью процессов зрительного анализатора (Э.Г. Вацуро, 1945; Э.Г. Вацуро, М.С. Ко-лесников, 1948). Такое заключение основывалось на результатах изменения мест зрительного и слухового раздражителей в стереотипе, а также на данных относительно переделки условных рефлексов, выработанных на световые и звуковые сигналы. В аналогичном исследовании, проведенном на обезьянах (Э.Г. Вацуро, 1947), было показано существование у них обратного соотношения: подвижность процессов в зрительном анализаторе оказалась более высокой, чем в слуховом.
В одной из работ Э.Г. Вацуро (1947) была отмечена у низших обезьян неуравновешенность в сторону преобладания возбуждения в слуховом анализаторе сравнительно со зрительным, а в другой (Э.Г. Вацуро, М.С. Колесников, 1948) указано на различия у этих обезьян в работоспособности корковых элементов зрительного и слухового анализаторов (в пользу последнего) и соответственно на различную «толерантность» этих анализаторов к кофеину.
На основании всех этих наблюдений Э.Г. Вацуро (1949) сформулировал положение о «ведущей афферентации», заключающееся в том, что разные анализаторы в зависимости от совершенства их функциональных свойств играют различную роль в организации поведения животного.
Напомним, однако, что эти различия имеют общевидовой характер, отражают особенности данного вида в целом, будучи, по-видимому, определены в филогенезе спецификой роли каждого анализатора в приспособлении организма данного вида к требованиям среды. Поэтому они, конечно, выпадают из контекста проблематики свойств нервной системы. Но нельзя, разумеется, исключить возможности того, что у отдельных особей отношение, «нормальное» для вида в целом, нарушается и соотношение свойств анализаторов в силу тех или иных причин приобретает индивидуальный характер. У человека никакой из основных анализаторов не играет специфически ведущей роли, поскольку главное регулирующее значение в процессе его эволюции приобретают факторы небиологического характера; но именно по этой причине у человека открываются большие, чем у животных, возможности интраиндивидуальных вариаций свойств отдельных анализаторов – вариаций, обусловленных, вероятно, в основном врожденными или унаследованными особенностями морфологической организации соответствующих кортикальных областей, но в какой-то мере, возможно, и условиями онтогенетического развития.
«То, что у животных было признаком вида, стало у человека признаком индивидуальности. В этом – одно из своеобразий человека. Особенно резко «ведущая афферентация» должна выступать у людей, основная жизненная деятельность которых способствует развитию относительного преобладания первой сигнальной системы: музыкантов, живописцев, спортсменов и др.» (Б.М. Теплов, 1956, с. 101).
Какие существуют фактические указания на то, что у человека 321 возможны межанализаторные различия в уровне тех или иных свойств нервной системы?
Косвенные свидетельства в пользу существования таких различий дают многочисленные психологические наблюдения над индивидуальными особенностями памяти («типы памяти» – зрительный, слуховой, двигательный), представлений, творческого воображения и т. д. Эти и подобные психологические явления – пока загадка с точки зрения их физиологической обусловленности. Представляя собой один из аспектов проблемы «специальной одаренности», они, как можно полагать, имеют прямое отношение к проблеме индивидуальных особенностей функционирования анализаторов. В связи с этим кажется весьма основательной гипотеза Б.М. Теплова о том, что «если общие типологические свойства определяют темперамент человека, то частные свойства имеют важнейшее значение при изучении специальных способностей» (1956, с. 102). Вопрос этот, однако, до сих пор не был подвергнут экспериментальному исследованию.
Столь же косвенный материал в пользу наличия межанализаторных различий содержится в некоторых разделах психофизиологии органов чувств. М. Вертхеймер (М. Wertheimer, 1955) показал, что флуктуации абсолютных зрительных и слуховых порогов, всегда в норме имеющие место, происходят практически независимо одни от других. Этот вывод находит подтверждение и в данных других авторов, упоминаемых М. Вертхеймером. Все эти данные можно толковать как свидетельство разнонаправленных колебаний возбудимости корковых концов соответствующих анализаторов, и тогда предположение о существовании межанализаторных различий получат определенную поддержку. Однако эти флуктуации можно понять и как следствие явлений чисто периферического, рецепторного характера. Вертхеймер именно на основании несовпадения сдвигов чувствительности разных модальностей считает причиной этих сдвигов процессы, происходящие в рецепторах. Однако, как будет видно из дальнейшего, определенную степень функциональной независимости могут получать в отдельных случаях и корковые отделы анализаторов.
Более определенный материал по проблеме межанализаторных различий могли бы принести прямые измерения связи между показателями чувствительности различных анализаторов. Известно, что диапазон индивидуальных колебаний показателей чувствительности в пределах нормы, по крайней мере, в зрительном и слуховом анализаторах составляет около 30 дБ (С.И. Вавилов, 1956; Н.И. Пинегин, 1946; Б.Е. Шейвехман и др., 1956). Идут ли эти различия параллельно по разным анализаторам, или они, подобно разнонаправленным колебаниям порогов, могут идти и в противоположных направлениях? Ответ на этот вопрос означал бы также ответ и на вопрос относительно возможности межанализаторных различий по уровню силы нервных клеток. К сожалению, в литературе мы такого ответа не обнаружили. Между тем в работе по исследованию связи между чувствительностью и силой нервной системы мы столкнулись с фактами, которые нельзя было расценить иначе, как свидетельство в пользу возможности «разведения» анализаторов в отношении их основных функциональных характеристик.
Определяя первоначально силу нервной системы с помощью угашения с подкреплением условной реакции, выработанной на звуковой раздражитель, а также с помощью индукционной методики и сопоставляя полученные показатели с цифрами зрительной чувствительности, мы встретились со случаями явного несовпадения данных первой и второй методик, а также со случаями, обратными предполагаемой зависимости между силой (по угашению с подкреплением) и абсолютными порогами. Возникло предположение, что эти случаи обусловлены тем, что измерения чувствительности (а в индукционной методике – и силы) относятся к зрительному анализатору, а угашение с подкреплением, использующее звуковой условный раздражитель, – к слуховому анализатору. Действительно, использовав при угашении с подкреплением зрительный условный стимул, а кроме того, перейдя к измерению абсолютных слуховых порогов и сопоставляя данные, относящиеся к одному и тому же анализатору, мы случаев несовпадения встретили уже гораздо меньше (см. об этом гл. VIII). Располагая группой количественных индикаторов силы нервных клеток, относящихся к разным анализаторам, мы могли теперь сравнить силовые характеристики анализаторов как сопоставлением непосредственных индикаторов силы, так и путем сравнения показателей абсолютной чувствительности (В.Д. Небылицын, 1957 а).
Подсчет коэффициента корреляции рангов между показателями чувствительности дал величину его, равную 0,263. Таким образом, корреляция между порогами, хотя и положительная, свидетельствует о наличии в сопоставляемых рядах значительного числа несовпадений, обусловливающих низкую величину связи.
Но если существуют межанализаторные различия по уровню чувствительности, то, поскольку чувствительность связана с силой, должны существовать и межанализаторные различия в уровне силы нервных клеток. Нужно допустить, следовательно, существование силы как парциального свойства нервной системы, характеризующего функцию отдельных зон и областей головного мозга. Мы полагаем, что наша работа доставила факты, определенно указывающие на такую возможность.
При сравнении у 25 испытуемых суммарных силовых характеристик зрительного и слухового анализаторов было обнаружено, что в большинстве случаев можно говорить о более или менее полном соответствии этих характеристик. Из 25 испытуемых такое соответствие констатировано у 18 человек, что составляет 72 % всех случаев. Однако наряду с этим были отмечены 7 случаев сравнительно отчетливых расхождений между силовыми показателями, из которых три случая, на наш взгляд, являются совершенно недвусмысленными. Рассмотрим эти три случая подробнее.
Испытуемый К.В. занимает второе место по уровню зрительной чувствительности. Все три способа определения силы нервных клеток в зрительном анализаторе (кроме упомянутых угашения с подкреплением и индукционной методики, еще и влияние кофеина на чувствительность) ясно показали слабость этих клеток (рис. 29 и 67): при испытании индукционной методики были отмечены сильные изменения концентрации в очаге дополнительного раздражителя под влиянием каждой из трех доз кофеина; угашение с подкреплением в зрительном анализаторе привело к снижению условной реакции и полному ее исчезновению в опыте с кофеином; прямое влияние кофеина на пороги (доза 0,3 г) выразилось в первоначальном снижении чувствительности, а затем в ее росте на 270 %. В то же время по слуховой чувствительности этот испытуемый занимает 24‑е место; проба на угашение с подкреплением при звуковом условном раздражителе приводит к увеличению реакции: влияние кофеина на слуховую чувствительность составляет 35 % повышения при максимальной дозе 0,3 г. Эти данные свидетельствуют о высоком уровне силы нервных клеток в слуховом анализаторе.
Испытуемая Р. З. по абсолютной зрительной чувствительности делит 20–21‑е места, испытание по индукционной методике не выявило какого-либо существенного влияния кофеина на характер первоначальной индукционной кривой; при угашении с подкреплением величина условной реакции на зрительный стимул почти не меняется; при максимальной дозе кофеина происходит некоторое повышение зрительной чувствительности (рис. 68). По слуховой чувствительности испытуемая Р. 3. занимает место в верхней половине ряда – 10‑е; проба на угашение с подкреплением приводит к снижению реакции, особенно в опыте с кофеином; прямое действие кофеина на слуховую чувствительность выражается в ее снижении на 40 % как при средней, так и при максимальной дозе. Таким образом, расхождения в уровне силы между зрительным и слуховым анализаторами у испытуемой Р. 3. направлены по сравнению с испытуемым К.В. в противоположную сторону: если К.В. обладает сильным слуховым и слабым зрительным анализаторами, то Р.З. – наоборот: слабым слуховым и сильным зрительным.
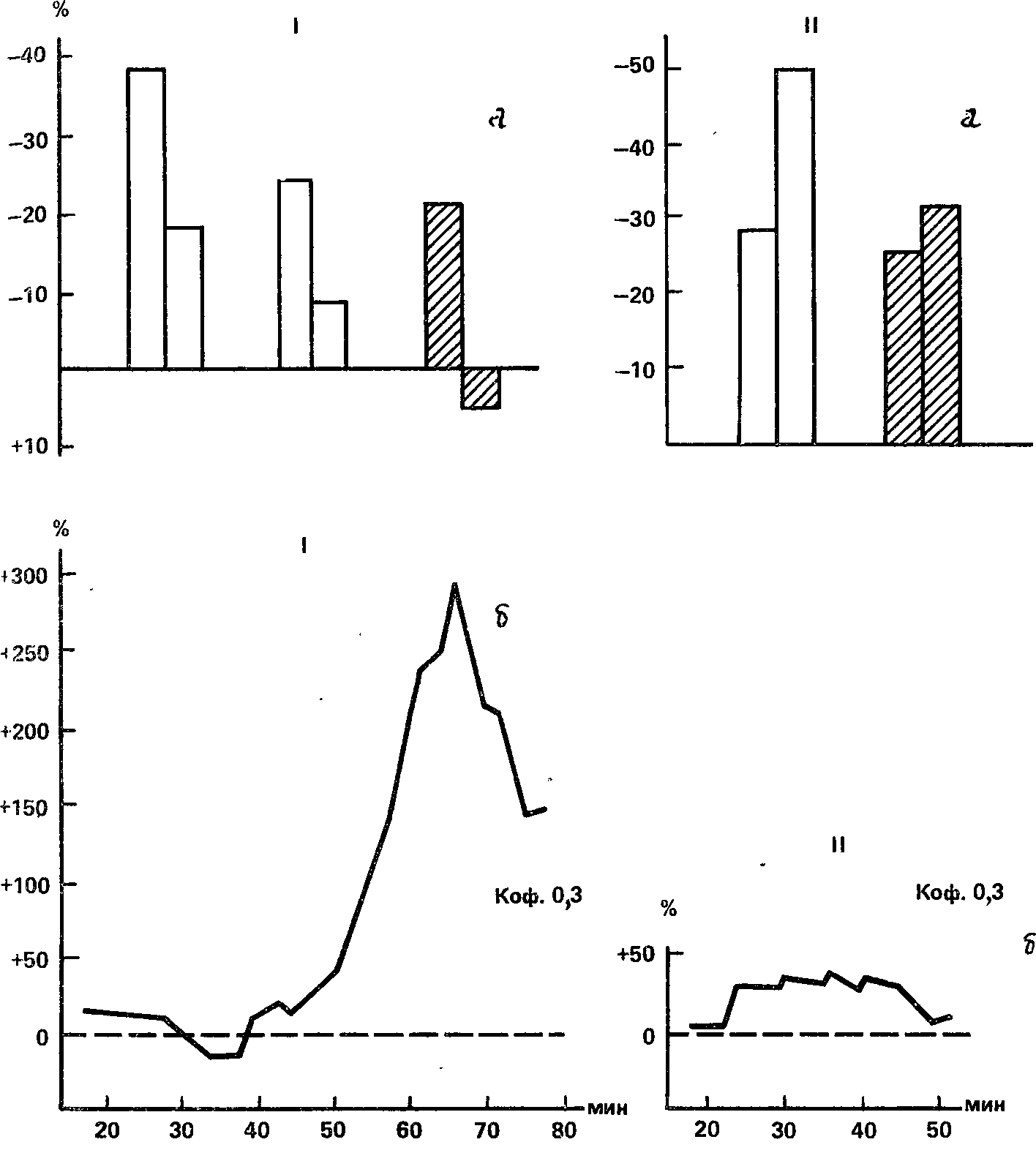
Рис. 67. Результаты определения силы нервной системы методиками, относящимися к зрительному (I) и слуховому (II) анализаторам, у исп. К.В. Обозначения: а – угашение с подкреплением, б – влияние кофеина на абсолютную чувствительностью (В.Д. Небылицын, 1957а).
Испытуемая Ч.И. имеет низкую зрительную чувствительность (19‑е место в группе), которая не изменяется при действии кофеина (рис. 69). Испытание по индукционной методике дало результаты, типичные для «сильных» испытуемых. Фотохимический условный рефлекс на световой сигнал у этой испытуемой не удалось выработать, и поэтому угашение с подкреплением не могло быть осуществлено. Что же касается слухового анализатора, то Ч.И. обладает очень высокой чувствительностью, занимая первое место в группе. Угашение с подкреплением при звуковом условном сигнале привело к глубокому падению реакции, так что условный раздражитель вызвал даже реакцию «обратного знака». Слуховая чувствительность под влиянием кофеина падает, при больших дозах на 20–23 %. Эти данные говорят об относительно высокой силе клеток зрительного анализатора и слабости – слухового, что сходно с характеристикой исп. Р. З.
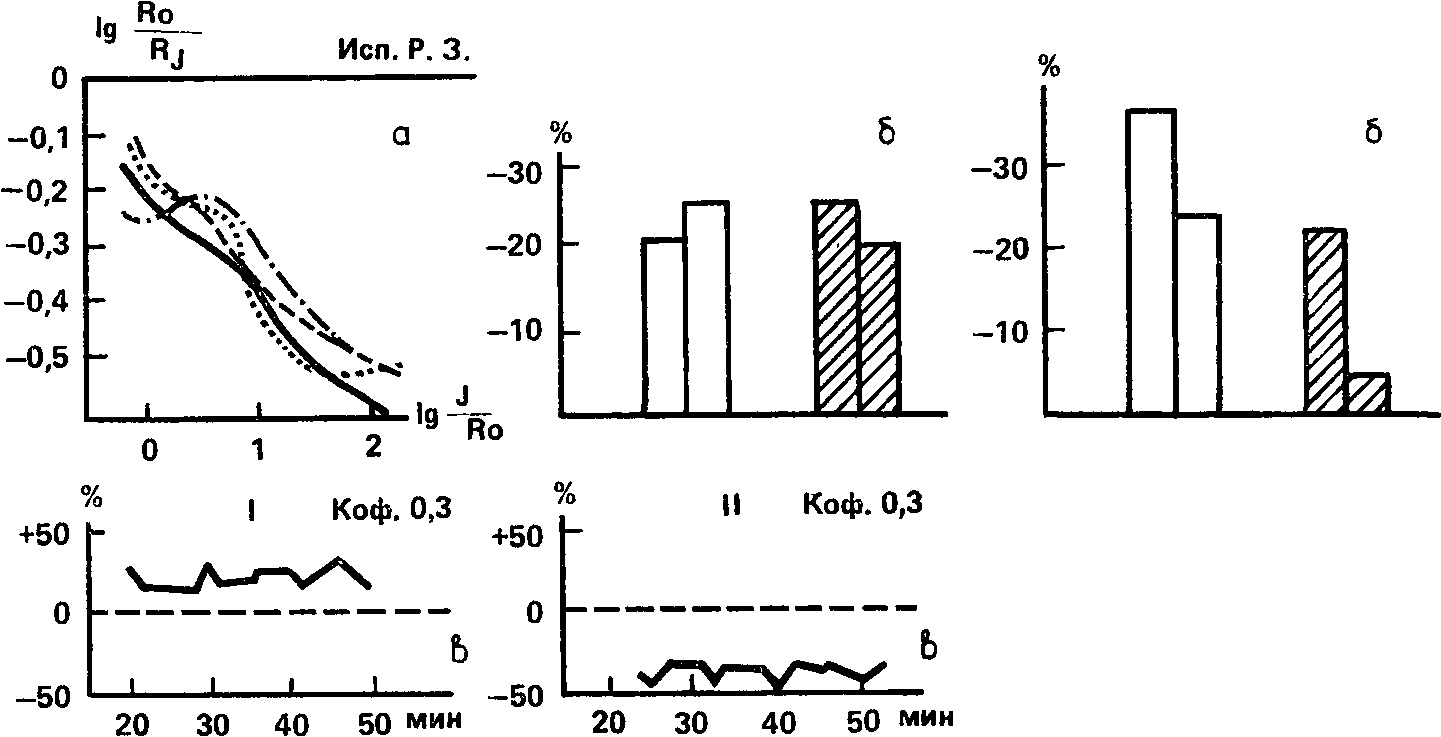
Рис. 68. Результаты определения силы нервной системы методиками, относящимися к зрительному (I) и слуховому (II) анализаторам, у исп. Р 3. Обозначения: а – индукционная методика, б – угашение с подкреплением, в-влияние кофеина на абсолютную чувствительность (В.Д. Небылицын, 1957а).
Если добавить к материалам этих трех испытуемых несколько менее определенные данные четырех других, то нужно будет заключить, что существуют такие индивиды, по отношению к которым термин «сила нервной системы» должен употребляться с указанием на то, какие анализаторы он характеризует. При этом подобные случаи не являются такими уж редкими. В нашем материале они составили около одной четверти, если взять всех семерых испытуемых, или 12 %, если ограничиться тремя наиболее ясными случаями расхождений.
Как видно из изложенного, основным индикатором силы корковых клеток служил в этой работе эффект действия кофеина, применявшегося во всех силовых методиках. Может возникнуть вопрос: поскольку опыты с воздействием кофеина проводились в разные опытные дни, не объясняются ли полученные различия, в частности в реакциях анализаторов при измерении непосредственного влияния кофеина на чувствительность, различиями в функциональном состоянии испытуемого?
Чтобы ответить на этот вопрос, с тремя охарактеризованными выше испытуемыми были поставлены специальные опыты, в которых измерение зрительной и слуховой чувствительности производилось в течение одного и того же промежутка времени, поочередно, с интервалом 1 – 2 мин. Результаты этого контрольного эксперимента совпали с теми, которые были получены в основных сериях (рис. 70): удалось наблюдать «сильную» реакцию одного анализатора при одновременной «слабой» реакции другого, с сохранением того же направления межанализаторных различий, которое было зарегистрировано ранее. Этот опыт позволяет заключить, что примененные методики вскрывают действительно устойчивые особенности функции анализаторов, представляющие их конституциональные свойства.
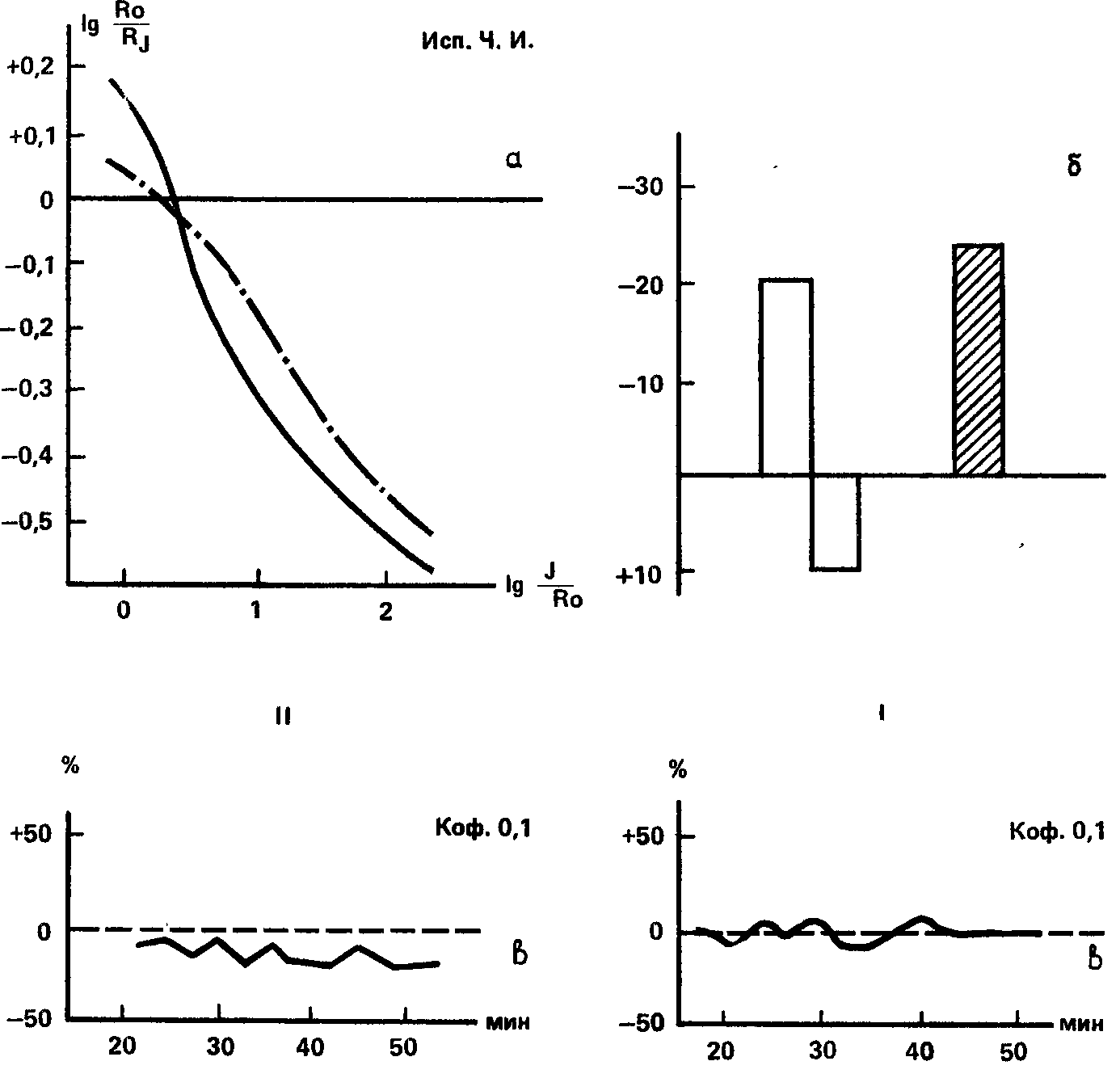
Рис. 69. Результаты определения силы нервной системы методиками, относящимися к зрительному (I) и слуховому (II) анализаторам, у исп. Ч.И. Обозначения те же, что на рис. 68 (В.Д. Небылицын, 1957а).
Весьма существенно то обстоятельство, что трем отмеченным наиболее резким случаям расхождений по уровню силы сопутствуют столь же резкие расхождения и по уровню абсолютной чувствительности. Следовательно, установленное ранее обратное соотношение между чувствительностью и силой нервной системы остается действительным и в том случае, когда понятие силы нервной системы должно рассматриваться в более узком аспекте, применительно к деятельности отдельного анализатора.
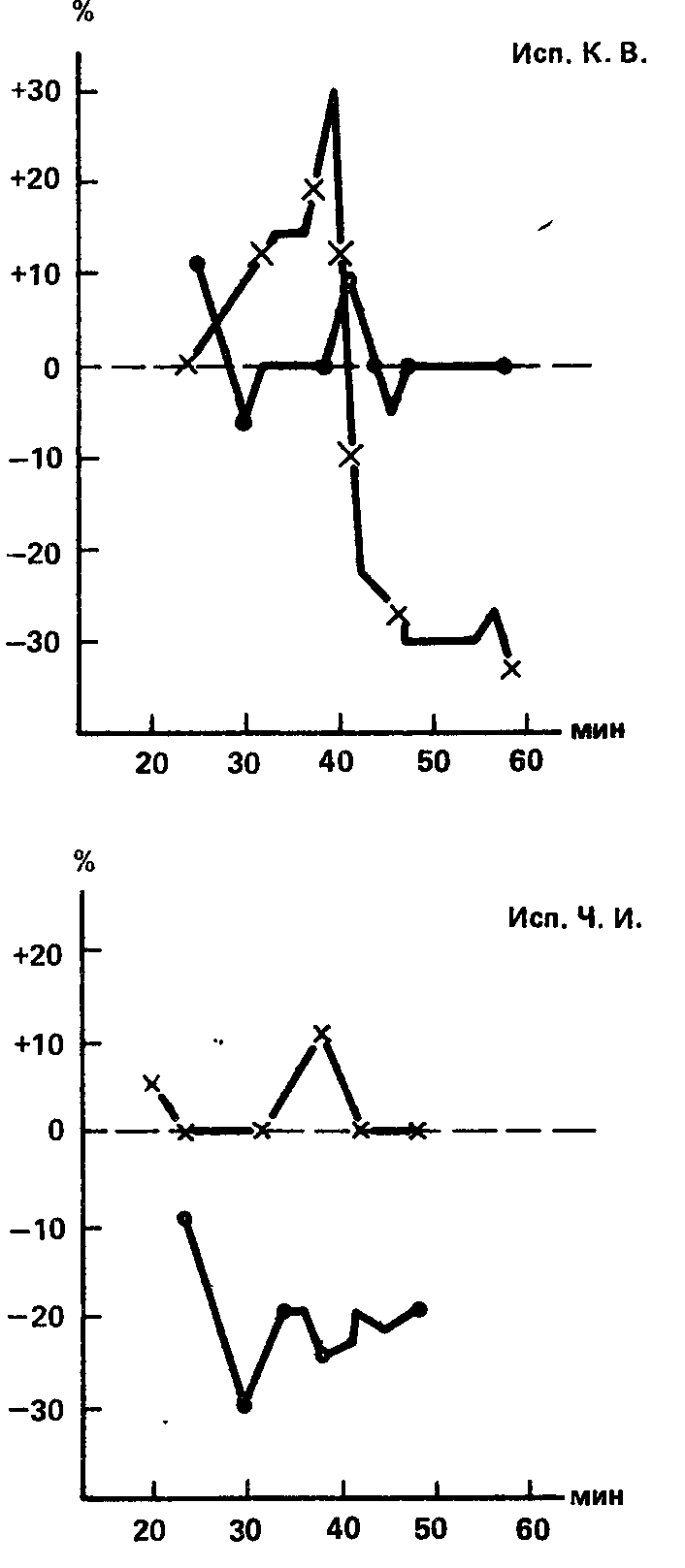
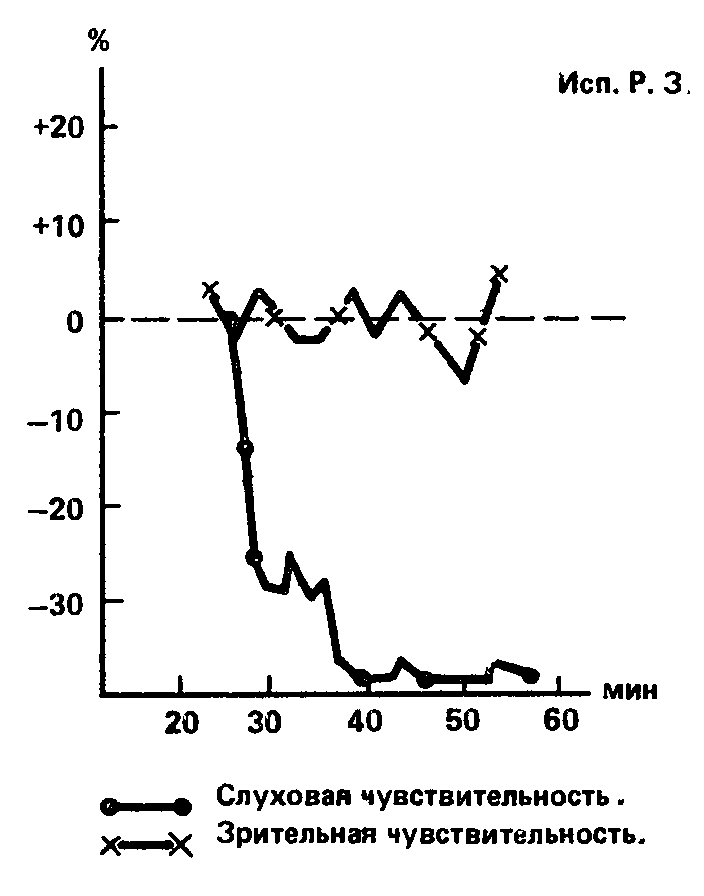
Рис. 70. Влияние 0,1 г чистого кофеина на слуховую и зрительную чувствительность в опытах с одновременным измерением порогов у трех испытуемых с межанализаторными различиями по уровню силы нервных клеток.
Ось абсцисс – время (в мин); ось ирдинат – изменения чувствительности (в %) по отношению к «фону» (В.Д. Небылицын, 1957а).
Анализ экспериментальных материалов, относящихся к проблеме парциальных проявлений силы нервной системы, позволил нам в свое время (В.Д. Небылицын, 1957а) прийти к одному методически существенному заключению. Оно касалось следующей проблемы: функцию какого анализатора исследует условный рефлекс, выработанный путем сочетания раздражителей разных сенсорных модальностей? И более конкретно: силу нервных клеток какого анализатора определяет угашение с подкреплением условного фотохимического рефлекса, выработанного при сочетаниях «звук плюс свет», – зрительного или слухового? Ответ на этот вопрос был дан, как уже говорилось, путем выработки условной фотохимической реакции на световой условный раздражитель, возможность чего была ранее показана Э.А. Голубевой (1955) и И.В. Равич-Щербо (1956), и сравнения двух видов угашения с подкреплением – при звуковом и световом условных сигналах – с другими индикаторами силы нервных клеток.
Результаты этого сравнения с определенностью указали на то, что угашение с подкреплением измеряет силу нервных клеток того анализатора, к которому обращено действие условного раздражителя, а не того, который принимает энергию подкрепления. Об этом говорят данные тех трех испытуемых, которые были разобраны выше. Некоторые литературные материалы также говорят в пользу этого предположения (З.А. Барсукова, 1956; И.И. Чинка, 1956). Ему, как мы полагаем, не противоречат и весьма обоснованные взгляды, развиваемые Э.А. Асратяном (1961) и его сотрудниками (Ф.К. Даурова, 1962), согласно которым запредельное (как и условное) торможение возникает первично не в афферентных клетках условного Сигнала и не в центре безусловного раздражителя, а в промежуточных нейронах, составляющих саму дугу условной связи: ведь, по крайней мере, начальная часть этой дуги включена в состав того анализатора, к которому адресуется энергия условного стимула.
Во всяком случае, представление о зависимости силовой характеристики, получаемой при помощи угашения с подкреплением, от модальности условного раздражителя вошло в практику экспериментальной работы лаборатории психофизиологии и было впоследствии неоднократно апробировано в ряде исследований, затрагивавших проблему межанализаторных различий.
Проблема эта была впоследствии вторично подвергнута специальному исследованию Ф.В. Ипполитовым (1964); с ней регулярно сталкивались также все авторы, одновременно применявшие в своей работе приемы определения силы нервных клеток, адресованные к разным анализаторам (собственно к зрительному и слуховому). Так, в работе Л.Б. Ермолаевой-Томиной (1959) из 32 испытуемых, исследованных двумя этими разновидностями методик, было обнаружено четверо индивидов (т. е. 12,5 %) с противоположным уровнем силы нервных клеток в зрительном и слуховом анализаторах, причем заметим, у двух из них несовпадение шло в одном направлении, а у двух других – в обратном.
В коллективной работе четырех авторов (В.И. Рождественская и др., 1960), проведенной на 40 испытуемых, наличие межанализаторных различий по уровню силы было установлено у 7 испытуемых (17,5 %), однако распределение их по направлению различий было довольно односторонним: шестеро из них имели сильные клетки зрительного анализатора и слабые – слухового и только один – наоборот. Интересно, что коэффициент корреляции рангов между зрительной и слуховой чувствительностью оказался, по данным этой работы, равен 0,34 (р < 0,05), т. е. довольно близок к тому, который был найден нами раньше. Другие корреляции между аналогичными, но относящимися к разным анализаторам испытаниями тоже были не очень высоки. Так, корреляция между двумя разновидностями угашения с подкреплением составила 0,41, что, хотя и значимо на 1 %-ном уровне вероятности, все же указывает на существование значительного «случайного» (по отношению к общему фактору силы) варьирования, а корреляция между двумя вариантами влияния кофеина на чувствительность была всего лишь 0,22. Это, однако, не помешало силовым индикаторам как зрительного, так и слухового анализатора при факторной обработке матрицы интеркорреляций войти в один и тот же фактор, идентифицированный как «сила нервной системы» в целом.
Определенные указания на возможность различий по уровню силы между зрительным и слуховым анализаторами содержатся также в работе З.Г.Туровской (1963б). Здесь, правда, индикатором силы слухового анализатора служил только один показатель – слуховая чувствительность (корреляция которой со зрительной была, кстати, 0,27, что снова весьма близко к прежним данным), однако автор имел возможность сравнить результаты некоторых своих испытуемых с данными по угашению с подкреплением, проведенному на тех же испытуемых В.И. Рождественской. Оказалось, что один из испытуемых, показавший высокую силу клеток зрительного анализатора (при испытании индукционной методикой) и в то же время имевший высокие показатели слуховой чувствительности, по данным В.И. Рождественской, действительно, обладает слабостью нервных клеток слухового анализатора. У двух других испытуемых соотношение имело противоположный характер.
Наконец, еще одна группа данных, на которую следует указать в связи с обсуждаемой проблемой, содержится в работе И.О. Майера (1963), применившего с целью испытания силы нервной системы по отношению к возбуждению у детей продление действия положительных условных раздражителей (методика речевого подкрепления). Автором было найдено, что у 33 детей из 57 (58 %) звуковой и световой сигналы дают совпадающий эффект, а у 24 результаты расходятся, что дало автору основания сделать вывод о различной силе нервных клеток соответствующих анализаторов у детей последней группы. В сообщении И.О. Майера привлекает внимание высокий процент случаев межанализаторных различий: он примерно в 2 раза больше того, который наблюдался в упомянутых выше работах сотрудников лаборатории Б.М. Теплова. Возможно, это явление связано со сложным характером силового испытания, примененного И.О. Майером. Хотя автор полагает, что этим испытанием определяется функциональная выносливость нервных клеток, есть все основания думать, что им, кроме того, испытывается и динамичность тормозного процесса (быстрота развития торможения при неподкреплении). Таким образом, межанализаторные различия, вскрываемые продлением действия условного сигнала, – это, видимо, различия сразу по двум факторам деятельности нервной системы. Естественно, что процент испытуемых с расхождениями характеристик анализаторов значительно повышается, если о нем судить по результатам этого комплексного испытания.
Так или иначе реальность феномена парциальности в его условно-афферентном проявлении вряд ли может сейчас вызывать сомнения. Существование индивидов с интраиндивидуальными вариациями свойств нервной системы, – во всяком случае, силы нервных клеток – представляется сейчас доказанным фактом. Тем не менее среди исследователей нет единого мнения по этому вопросу. Так, В.С. Мерлин (1964), ссылаясь на некоторые работы сотрудников лаборатории Б.М. Теплова, утверждает, что «эти данные отнюдь не говорят о том, что разные анализаторы могут обладать разными свойствами общего типа» (1964, с. 90). В обоснование своей точки зрения В.С. Мерлин указывает, что результаты испытания силы нервных клеток зависят от физической интенсивности стимула, а в приведенных им работах надлежащего учета этого фактора при сопоставлении звуковых и световых раздражителей не было.
Возражение кажется резонным, однако оно имело бы смысл только в том случае, если бы мы в каждом данном сопоставлении (скажем, двух разновидностей угашения с подкреплением) находили примеры только одного направления различий между анализаторами, например только слабости зрительного анализатора при высокой силе слухового или наоборот. В этом случае мы действительно могли бы полагать, что один из условных раздражителей физически слишком интенсивен по сравнению с другим. Однако в каждой из упомянутых выше работ встречались случаи и того и другого направлений межанализаторных различий. Из зарегистрированных к настоящему времени в работах сотрудников Б.М. Теплова 17 случаев отчетливых расхождений по силе нервных клеток в 11 случаях имела место слабость в слуховом анализаторе в сочетании с высокой силой в зрительном анализаторе, а в 6 случаях – обратное соотношение силовых показателей. Отсюда следует, что фактор физической интенсивности условного стимула, который в принципе, конечно, имеет большое значение для силовой характеристики, в данных работах не играл существенной роли.
Другое возражение В.С. Мерлина состоит в следующем: поскольку показатели силы зависят не столько от физической, сколько от физиологической силы раздражителя, а последняя зависит от возбудимости анализатора, то, «применяя объективно одни и те же раздражители у лиц, резко различающихся по сенсорной возбудимости данного анализатора, мы должны получить такие же результаты, как если бы применяли раздражители разной силы» (1964, с. 90–91). Отсюда следует, очевидно, что физиологическая сила раздражителей должна быть уравнена, т. е. стимулы должны браться в единицах порога. Едва ли, однако, такой подход будет эффективен в исследовании проблемы силы нервной системы: ведь изложенный в гл. VIII и IX материал ясно свидетельствует о том, что дифференцирование испытуемых по параметру силы тем и достигается, что им предъявляются раздражители различной физиологической эффективности (различной как раз ввиду разницы в их порогах). Другое дело – изучение таких свойств нервной системы, как подвижность или динамичность. Здесь, действительно, для того чтобы исключить влияние параметра чувствительности – силы, следует уравнивать раздражители по их физиологической эффективности. Но В.С. Мерлин имеет в виду именно силу нервной системы.
Мотивы возражений, выдвигаемых В.С. Мерлиным, достаточно ясны. В самом деле, если существуют – и не так уж редки – парциальные особенности свойств нервной системы, то не рассыпается ли само понятие свойства нервной системы как целостной устойчивой характеристики определенной стороны ее деятельности, не исчезает ли нейрофизиологическая основа общих свойств приспособительной деятельности и поведения индивида в целом? При ответе на эти вопросы следует, как мы полагаем, исходить из следующих соображений.
Во-первых, согласно экспериментальным данным, существенные межанализаторные различия (по крайней мере, между двумя основными анализаторами) имеют место в среднем лишь у 15–20 % выборки, подвергаемой исследованию. Таким образом, по отношению к подавляющему большинству (80–85 %) испытуемых проблема парциальности не возникает.
Во-вторых, у тех испытуемых, которые обнаруживают расхождения между анализаторами по уровню того или иного свойства, видимо, существует как их индивидуальная особенность тот самый «ведущий» анализатор, о котором говорится в цитированном выше высказывании Б.М. Теплова и доминирование которого накладывает отпечаток на проявления психической организации индивида. В случае наличия «ведущего» анализатора о свойствах нервной системы, возможно, и следует судить по результатам исследования этого анализатора.
В-третьих, при оценке «общих» свойств нервной системы теоретически возможен и принципиально иной подход по сравнению с тем, который полагает ответственными за параметры нервной системы качества только корковых клеток и процессов. Учитывая, что существуют инфракортикальные мозговые структуры с интегральным характером эффекта, обеспечивающие в неразрывном единстве с кортикальными образованиями общий тонус организма и самые общие динамические характеристики реагирования, можно предположить, что целостные параметры мозговой организации, «общие» свойства нервной системы определяются синтезом функциональных характеристик подкорки и коры, в котором далеко не последнюю роль играют свойства общеактивирующей неспецифической системы.
В одной из публикаций мы уже сделали попытку обосновать значение неспецифических ретикулярных образований как возможного мозгового субстрата динамичности процесса возбуждения (В.Д. Небылицын, 19646). Но, вероятно, роль этих образований в многомерной структуре свойств нервной системы отнюдь не исчерпывается обеспечением функции замыкания, она значительно шире. Укажем в подтверждение этой мысли хотя бы на данные Е. Цукермана (Е. Zuckermann, 1959; А. Крейндлер, 1962), нашедшего лучшее сохранение условных рефлексов при угашении с подкреплением в условиях стимуляции ретикулярных структур. На эту же мысль наводят материалы Т.С. Наумовой (1961), указывающей на основании обнаруженного ею сходства процессов в специфических и неспецифических системах каждого отдельного животного на возможность того, что «типологические особенности нервной системы животных определяются теми процессами, которые протекают как в коре головного мозга, так и в стволовых структурах», и, более конкретно, что «канву индивидуальных различий создает характер тех диффузных генерализованных процессов мозга, которые определяют целостную реакцию (со всеми ее компонентами) организма на безусловные раздражения» (1961, с. 139). Можно думать, что синдром каждого из параметров мозговой деятельности в существенной степени детерминирован качественными особенностями функции тех мозговых образований, которые имеют равное отношение ко всем корковым зонам и отделам и которые в единстве с последними составляют структурно-морфологическую основу «над-анализаторных» проявлений основных свойств нервной системы.
Разумеется, такая постановка вопроса имеет чисто гипотетический характер, однако она, как нам кажется, оправдана эволюцией наших знаний относительно роли субкортикальных образований в условнорефлекторной деятельности. Так или иначе возможность самого существования топографически обусловленных, зависящих от территориальной приуроченности испытания проявлений свойств нервной системы вряд ли может вызывать сомнения; такие проявления, как мы видели, обнаруживаются всякий раз, когда применяются соответствующие формы испытаний. Вопрос же о психофизиологическом смысле этого феномена еще ожидает своего разрешения.

