 2015-06-26
2015-06-26 1242
1242Первым этапом в регуляции экспрессии генов при водном дефиците является рецепция сигнала. Воспринятый клеткой сигнал приводит к включению систем сигнальной трансдукции и активированию белков, образующих цепь передачи сигнала. Имеются данные, свидетельствующие о том, что у растений рецепция сигнала при водном стрессе осуществляется аналогично тому, как это происходит у Е. coli и дрожжей. У этих организмов восприятие изменений осмотического давления наружной среды совершается с помощью сенсорной системы, состоящей из локализованного в мембране осмосенсора (сенсорной киназы) и находящегося в цитоплазме регулятора ответа (рис. 8.10).
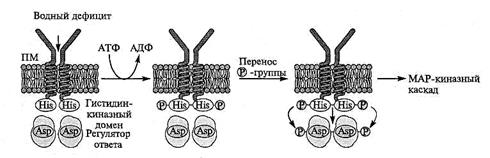
Рис. 8.10. Двухкомпонентная сенсорная система, воспринимающая изменения осмотического давления наружной среды (A.Trewavas, 2000).
Система состоит из локализованного в мембране осмосенсора и находящегося в цитозоле регулятора ответа. Сигнал возникает при изменении натяжения плазмалеммы в ответ на изменение осмотического давления наружной среды. При получении сигнала осмосенсор, подвергаясь автофосфорилированию, активируется. С гистидинового остатка молекулы осмосенсора фосфатная группа затем переносится на остаток аспарагиновой кислоты регулятора ответа. Фосфорилированная молекула регулятора ответа приводит к включению МАР-киназного пути передачи сигнала
Эта система воспринимает изменения в натяжении ПМ при водном дефиците. В ПМ дрожжей осмосенсор, трансмембранный белок Slu 1 p, при высокой осмолярности среды активируется путем автофосфорилирования, приводя затем к последовательному фосфорилированию белков цитоплазмы Ypd 1 и Ssk 1 p, которые образуют регулятор ответа.
МАР-киназный каскад. У дрожжей фосфорилированный белок Ssklp активирует различные пути передачи сигнала, в частности MAP(mitogen activated protein)-киназный каскад, состоящий из трех последовательно фосфорилируе-мых в присутствии АТФ протеинкиназ: Ssk 2 p (MAPKKK) — МАРКК-киназы, Pbs 2 p (MAPKK)— МАРК-киназы и Hog 1 p (МАРК) — МАР-киназы (рис. 8.11).
Активированная фосфорилированием МАР-киназа индуцирует экспрессию многих генов дрожжевой клетки при водном и солевом стрессах, в частности генов, кодирующих ферменты биосинтеза глицерина — главного осмолита у дрожжей. Гомологи некоторых компонентов МАР-киназного каскада дрожжевых клеток обнаружены у растений. Экспрессия генов этих компонен- тов индуцируется стрессовыми условиями. В частности, у A. thaliana ген МАР-киназы (АТМРКЗ) и ген МАРККК (АТМЕКК1) индуцируются засухой, низкими температурами и высокими концентрациями соли.
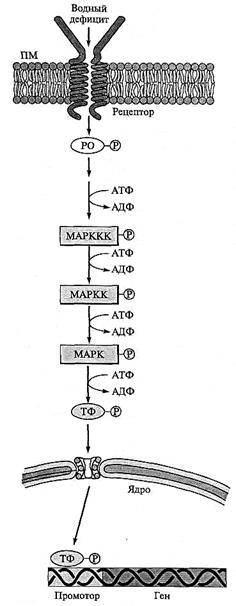
Рис. 8.11. МАР-киназный путь передачи сигнала (по A. Trewavas, 2000, с изменениями).
Фосфорилированные белки регулятора ответа (РО) активируют МАР-киназный каскад, состоящий из МАРККК (МАРКК-киназы), МАРКК (МАРК-киназы) и МАРК (МАР-киназы). Последняя осуществляет прямую регуляцию транскрипционных факторов (ТФ), вовлеченных в индукцию стрессовых белков при водном дефиците
Вторичные мессенджеры. В регуляции тургорного давления растительной клетки участвуют вторичные мессенджеры (рис. 8.12). Локализованный в мембране фосфа-тидилинозитол-4,5-дифосфат расщепляется активируемой при осмотическом стрессе фосфолипазой С, освобождая вторичный мессенджер инозитол-1,4,5-трифосфат. Последний индуцирует выход Са2+, другого вторичного мессенджера, из вакуоли и эндоплазматического ретикулума в цитозоль и соответственно концентрация Са2+ в нем возрастает. Концентрация Са2+ в цитозоле может увеличиваится также за счет его поступления в клетку через потенциалзависимые Са2+-каналы плазмалеммы при индуцируемой стрессором деполяризации этой мембраны. Возросшие концентрации Са2+ активируют Са2+/кальмодулинзависимые протеинкиназы (CDPK), что в свою очередь стимулирует биосинтез стрессовых белков.
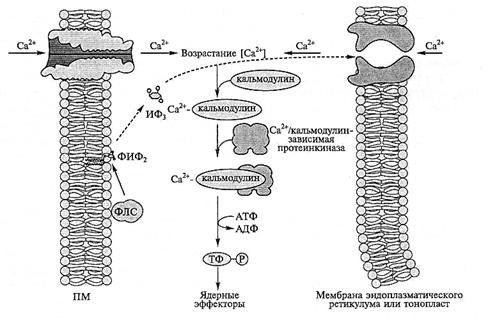
Рис. 8.12. Участие вторичных мессенджеров, инозитол-1,4,5-трифосфата и ионов Са2+ в путях передачи сигналов, приводящих к индукции стрессовых белков при водном дефиците:
ФЛС — фосфолипаза С; ФИФ2 — фосфатидилинозитол-4,5-дифосфат; ИФ3 — инозитол-1,4,5-трифосфат;
ТФ — транскрипционные факторы
АБК-зависимые и АБК-независимые гены. Важную роль в регуляции экспрессии генов при осмотическом стрессе играет гормон АБК. Экспрессия многих генов в условиях засухи, засоления или низких температур требует предварительного накопления в тканях эндогенной АБК, тогда как экспрессия других генов происходит без участия АБК. На основании этого и анализа действия АБК на экспрессию индуцируемых водным дефицитом генов в АБК-дефицитном (aba) и АБК-нечувствительном (аbi) мутантах A. thaliana был сделан вывод, что существуют АБК-зависимые и АБК-независимые пути передачи сигнала в ответной реакции растений на водный дефицит (рис. 8.13).
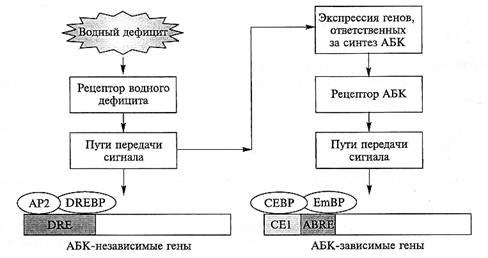
Рис. 8.13. Индукция АБК-зависимых и АБК-независимых генов водным дефицитом.
Промоторная область АБК-независимых генов содержит специфическую последовательность нуклеотидов DRE, которая связывается с транскрипционными факторами, белками DREBP и АР2. Промоторная область АБК-зависимых генов содержат нуклеотидную последовательность ABRE, которая связывается с транскрипционным фактором — белком EmBP. Помимо ABRE промоторная область АБК-зависимых генов включает сопрягающий элемент СЕ1, взаимодействующий с белком СЕВР
Несколько транскрипционных факторов и специфических нуклеотидных последовательностей в генах идентифицированы для обоих путей. АБК-индуцируемые гены содержат в промоторной области специфическую последовательность нуклеотидов, получившую сокращенное название ABRE (ABA-responsive element). ABRE функционирует как цис -элемент ДНК, вовлеченный в АБК-регулируемую экспрессию генов. Такого рода элемент был сначала идентифицирован в Em-гене пшеницы (Triticum aestivum), а затем обнаружен в
АБК-индуцируемых генах кукурузы (Zea mays), ячменя, риса, табака и арабидопсиса. Для индукции экспрессии генов необходимо, чтобы ABRE связался с белком EmBPl. В большинстве случаев ABRE достаточен для АБК-регулируемой экспрессии генов, но в некоторых генах ABRE ассоциирован с еще одной нуклеотидной последовательностью, которая называется сопрягающим элементом (СЕ). Например, в гене HVA22 ячменя сопрягающий элемент СЕ1 требуется для экспрессии наряду с ABRE. Для экспрессии таких генов требуется еще один ДНК-связывающий белок, специфичный к СЕ, — СЕВР (coupling element binding protein). ABRE, СЕ и оба белка EmBP1 и СЕВР образуют комплекс, индуцирующий экспрессию АБК-зависимого гена. Однако до настоящего времени не ясно, как АБК активирует EmBP1, вызывая его последующее взаимодействие с ABRE и инициацию транскрипции. В ряде генов, регулируемых с участием АБК, обнаружены другие цис- элементы, отличающиеся от ABRE. Индукция таких генов происходит при специфическом связывании этими элементами транскрипционных факторов MYC и MYB.
Промоторная область АБК-независимых генов содержит последовательность нуклеотидов DRE (drought-responsive element). DRE вовлечен в регуляцию генов, индуцируемых непосредственно водным дефицитом и не требующих АБК для экспрессии. Транскрипционные факторы для таких генов — DRE-связывающие белки DREBP и АР2 — обнаружены в ядерном экстракте из
А. thaliana.

