 2015-07-03
2015-07-03 6231
6231Раздел 5 Водный обмен растений
Молекулярная структура, физико-химические свойства воды и ее физиологическое значение в растении (окончание)
Термодинамические основы водного обмена растений.
Для описания закономерностей транспорта воды используют такие понятия, как химический потенциал воды, осмотическое давление, водный и матричный потенциалы.
Химический потенциал воды, активность воды
Любое вещество, в том числе вода, движется в направлении уменьшения ее свободной энергии, т.е. в направлении ее более низкого химического потенциала. Химический потенциал – это мера энергии, которую данное вещество использует на реакцию или движение. В соответствии с законами термодинамики, химический потенциал воды (μw) при изменяющемся давлении можно представить уравнением
_
μw=μоw+RTln a w + V w Р (1)
 где μw – стандартный химический потенциал воды (Дж/моль), R – газовая постоянная (8,314 Дж/моль∙К); Т – абсолютная температура; аw – активность воды; Vw – парциальный молярный объем воды; Р – давление.
где μw – стандартный химический потенциал воды (Дж/моль), R – газовая постоянная (8,314 Дж/моль∙К); Т – абсолютная температура; аw – активность воды; Vw – парциальный молярный объем воды; Р – давление.
Химический потенциал, как и другие виды потенциальной энергии, является относительной величиной, поэтому в выражение химического потенциала включен стандартный химический потенциал μоw. Поскольку при расчетах движущих сил транспорта воды используют разность химических потенциалов в двух рассматриваемых точках системы, а не их абсолютные значения, то μоw сокращаются при вычитании и не фигурируют в выражении градиента химического потенциала, определяющего направление перемещения воды.
Влияние концентрации воды в системе на ее химический потенциал представлено членом RTln a w. Активность воды аw характеризует ту реальную концентрацию, соответственно которой вода участвует в различных процессах. Активность чистой воды равна единице. Всякие межмолекулярные и иные связи уменьшают подвижность и рассеиваемость молекул и снижают активность воды. В растворе и в клетке активность воды всегда меньше единицы. Из уравнения (1) следует, что химический потенциал воды тем выше, чем больше ее активность (концентрация). Растворенные вещества снижают активность воды, так как уменьшают ее мольную долю в системе и соответственно концентрационную составляющую ее химического потенциала. Член VwP в уравнении отражает влияние давления на химический потенциал. Развивающееся в биологических системах давление по своей природе гидростатическое, по величине оно может быть как больше, так и меньше атмосферного. Примером гидростатического давления, больше атмосферного, является давление протопласта на клеточную стенку, называемое тургорным давлением. Пример гидростатического давления меньше атмосферного – давление в сосудах ксилемы при транспирации. Следует обратить внимание на то, что в физиологии растений при исследовании влияния давления на транспорт воды за нулевую точку отсчета принимается давление, равное атмосферному. В соответствии с этой шкалой гидростатическое давление протопласта на КС является величиной положительной, а гидростатическое давление в сосудах ксилемы при транспирации – отрицательной. Величина Vw – парциальный молярный объем воды – показывает, насколько увеличивается объем системы с увеличением в ней количества воды на 1 моль.
Член VwP в уравнении отражает влияние давления на химический потенциал. Развивающееся в биологических системах давление по своей природе гидростатическое, по величине оно может быть как больше, так и меньше атмосферного. Примером гидростатического давления, больше атмосферного, является давление протопласта на клеточную стенку, называемое тургорным давлением. Пример гидростатического давления меньше атмосферного – давление в сосудах ксилемы при транспирации. Следует обратить внимание на то, что в физиологии растений при исследовании влияния давления на транспорт воды за нулевую точку отсчета принимается давление, равное атмосферному. В соответствии с этой шкалой гидростатическое давление протопласта на КС является величиной положительной, а гидростатическое давление в сосудах ксилемы при транспирации – отрицательной. Величина Vw – парциальный молярный объем воды – показывает, насколько увеличивается объем системы с увеличением в ней количества воды на 1 моль.
Вода может поступать в клетки растений благодаря набуханию биоколлоидов, увеличению степени их гидратации. Такое поступление воды характерно для сухих семян, помещенных в воду. Однако главным способом поступления воды в живые клетки является ее осмотическое поглощение.
Осмосом называется прохождение растворителя в раствор, отделенный от него полупроницаемой мембраной (т. е. пропускающей растворитель, но не молекулы растворенных веществ).
В Пфеффер предложил осмометр, названный им «искусственной клеткой». Во внутреннюю полость пористого фарфорового сосуда наливался раствор желтой кровяной соли K4[Fe(CN)6], и сосуд помещался в раствор CuSО4. При взаимодействии этих веществ в порах сосуда образовывалась гелеобразная масса железистосинеродистой меди Cu2[Fe(CN)6], служившая полупроницаемой мембраной.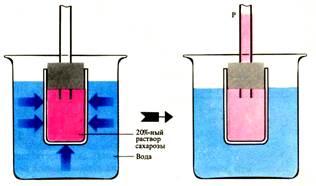 Демонстрация осмотического давления во внутреннем сосуде осмометра после его наполнения раствором сахарозы и погружения в чистую воду Раствор сахарозы отделен от воды мембраной, проницаемой для Н2О, но не для сахарозы. Р — гидростатическое давление
Демонстрация осмотического давления во внутреннем сосуде осмометра после его наполнения раствором сахарозы и погружения в чистую воду Раствор сахарозы отделен от воды мембраной, проницаемой для Н2О, но не для сахарозы. Р — гидростатическое давление Если раствор сахарозы в осмотической ячейке, пропускающей только воду, погрузить в чистую воду, то молекулы воды будут перемещаться через стенки ячейки в раствор сахарозы, где концентрация (активность) воды меньше, т. е. от высокого водного потенциала к более низкому. Растворенные вещества снижают активность воды в растворе и из ячейки воды выходит меньше, чем входит. Это и приводит к увеличению объема раствора сахарозы и подъему жидкости в трубке осмометра.
Поступление воды в такого рода осмотическую ячейку приводит к увеличению объема жидкости до тех пор, пока гидростатическое давление Р столба жидкости в монометрической трубке не повысится настолько, чтобы воспрепятствовать дальнейшему увеличению объема раствора. В достигнутом состоянии равновесия полупроницаемая мембрана в единицу времени пропускает одинаковые количества воды в обоих направлениях. Гидростатическое давление в этом случае соответствует потенциальному осмотическому давлению π*.Осмотическое давление – это давление, избыточное по отношению к атмосферному, которое необходимо приложить к раствору вещества, чтобы привести его в равновесие с чистым растворителем (или с раствором этого вещества иной концентрации), отделенным от него полупроницаемой мембраной. RT π = – –––– ln a w (2) Vw Водный потенциал Физиологическая активность растительной клетки зависит от термодинамического (энергетического) состояния воды, важнейшим показателем которой является водный потенциал. Разность химического потенциала воды в клетке и химического потенциала чистой воды (μ w– μо w), отнесенную к парциальному объему воды в клетке, называют водным потенциалом (ψв или ψ): μ w– μо w ψв = –––––––––– (3)
π = – –––– ln a w (2) Vw Водный потенциал Физиологическая активность растительной клетки зависит от термодинамического (энергетического) состояния воды, важнейшим показателем которой является водный потенциал. Разность химического потенциала воды в клетке и химического потенциала чистой воды (μ w– μо w), отнесенную к парциальному объему воды в клетке, называют водным потенциалом (ψв или ψ): μ w– μо w ψв = –––––––––– (3)  Vw Водный потенциал можно выразить соотношением: ψв = Р – π (4) Химический потенциал (μ) чистой воды всегда выше химического потенциала воды в клетке, поэтому величина водного потенциала всегда отрицательна. Водный потенциал растений выражается в атмосферах, барах или паскалях (1атм=1,013бар=105Па).Выражение водного потенциала для паров воды в воздухе имеет вид:
Vw Водный потенциал можно выразить соотношением: ψв = Р – π (4) Химический потенциал (μ) чистой воды всегда выше химического потенциала воды в клетке, поэтому величина водного потенциала всегда отрицательна. Водный потенциал растений выражается в атмосферах, барах или паскалях (1атм=1,013бар=105Па).Выражение водного потенциала для паров воды в воздухе имеет вид:  μ w– μо w RT RT относительная влажность ψв = –––––––––– = ––––– ln a w=––– ln –––––––––––––––––––––––– (5) Vw Vw Vw 100 В соответствии с уравнением (5) при относительной влажности, равной 100 %, водный потенциал паров воды в воздухе будет равен 0. Снижение относительной влажности всего лишь на 1 % приводит к уменьшению ψв до -1,36 МПа. Средние значения водного потенциала зеленых листьев, например овощных растений, составляют -0,5 МПа. Согласно уравнению (5), такое значение ψв соответствует относительной влажности 99,6 %. Даже во время проливного дождя влажность воздуха редко превышает 99 %. Следовательно, листья, водный потенциал которых составляет -0,5 МПа, постоянно теряют воду. В листьях большинства наземных растений водный потенциал колеблется от -0,4 до -0,3 МПа. У некоторых растений пустынь водный потенциал растений может снижаться до -5,0 МПа. Составляющие водного потенциала: осмотический потенциал, матричный потенциал, потенциал давления (гидростатический потенциал) В водном обмене растений большое значение имеет взаимодействие воды с биомолекулами. Такое взаимодействие осуществляется как внутри клеток, так и в апопласте и происходит в основном за счет образования водородных связей между молекулами воды и биополимерами. Водородные связи постоянно распадаются и образуются вновь. Связывание части молекул воды функциональными группами биополимеров понижает ее термодинамическую активность. Таким образом, биомолекулы наряду с низкомолекулярными растворенными веществами участвуют в снижении активности воды и вносят вклад в снижение ее химического потенциала. В связи с этим фактически измеряемое в экспериментах осмотическое давление в клетках, равное
μ w– μо w RT RT относительная влажность ψв = –––––––––– = ––––– ln a w=––– ln –––––––––––––––––––––––– (5) Vw Vw Vw 100 В соответствии с уравнением (5) при относительной влажности, равной 100 %, водный потенциал паров воды в воздухе будет равен 0. Снижение относительной влажности всего лишь на 1 % приводит к уменьшению ψв до -1,36 МПа. Средние значения водного потенциала зеленых листьев, например овощных растений, составляют -0,5 МПа. Согласно уравнению (5), такое значение ψв соответствует относительной влажности 99,6 %. Даже во время проливного дождя влажность воздуха редко превышает 99 %. Следовательно, листья, водный потенциал которых составляет -0,5 МПа, постоянно теряют воду. В листьях большинства наземных растений водный потенциал колеблется от -0,4 до -0,3 МПа. У некоторых растений пустынь водный потенциал растений может снижаться до -5,0 МПа. Составляющие водного потенциала: осмотический потенциал, матричный потенциал, потенциал давления (гидростатический потенциал) В водном обмене растений большое значение имеет взаимодействие воды с биомолекулами. Такое взаимодействие осуществляется как внутри клеток, так и в апопласте и происходит в основном за счет образования водородных связей между молекулами воды и биополимерами. Водородные связи постоянно распадаются и образуются вновь. Связывание части молекул воды функциональными группами биополимеров понижает ее термодинамическую активность. Таким образом, биомолекулы наряду с низкомолекулярными растворенными веществами участвуют в снижении активности воды и вносят вклад в снижение ее химического потенциала. В связи с этим фактически измеряемое в экспериментах осмотическое давление в клетках, равное 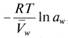 , можно представить в виде суммы π = πs + τ,где πs – осмотическое давление, вызываемое присутствием низкомолекулярных растворенных веществ; τ — матричное давление, являющееся результатом взаимодействия воды с биополимерами. Принимая во внимание наличие двух составляющих осмотического давления, мы можем переписать выражение ψв = Р – π следующим образом: ψв = Р – πs – τ. Часто в физиологии водного обмена водный потенциал представляют в виде сумммы потенциалов: ψв = ψp + ψs + ψm, где ψm – матричный потенциал, равный - τ ψp – гидростатический (потенциал давления), равный Р; ψs – осмотический потенциал давления, равный -πs.
, можно представить в виде суммы π = πs + τ,где πs – осмотическое давление, вызываемое присутствием низкомолекулярных растворенных веществ; τ — матричное давление, являющееся результатом взаимодействия воды с биополимерами. Принимая во внимание наличие двух составляющих осмотического давления, мы можем переписать выражение ψв = Р – π следующим образом: ψв = Р – πs – τ. Часто в физиологии водного обмена водный потенциал представляют в виде сумммы потенциалов: ψв = ψp + ψs + ψm, где ψm – матричный потенциал, равный - τ ψp – гидростатический (потенциал давления), равный Р; ψs – осмотический потенциал давления, равный -πs. Направление движения воды из наружной среды в клетку или обратно будет определяться направлением градиента водного потенциала. Условием поступления воды в клетку является более высокий водный потенциал наружной среды, чем водный потенциал клетки. Направление движения воды в системе целого растения также определяется направлением градиента водного потенциала. Осмотическая концентрация вакуолярного сока для клеток корней составляет 0,3 – 1,2 МПа, а для клеток надземных органов – 1,0-2,6 МПа. Это обусловливает существование вертикального градиента осмотической концентрации и сосущей силы от корней к листьям. Очень высоко осмотическое давление в клетках галофитов, растущих в условиях засоления: оно достигает 15 МПа.

