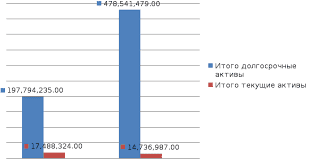
2015-08-21
2015-08-21 740
740сомнения, соответствующим инстинктом. Затем промежутки между скоплениями заполняются и в конечном счете распределение по поверхности становится равномерным. Все же плотность заселения плиты составляет лишь 35—55 особей на 1 см2, тогда как циприсовидные личинки эль-миниусов, длина которых не превышает 0,5 мм, а ширина еще меньше, способны заселять ровные поверхности с плотностью до 230 особей на 1 см2 даже в естественных условиях. При тех плотностях, при которых вступает в действие механизм, регулирующий распределение по поверхности, еще остается много места, так что запоздалым пришельцам есть где прикрепиться и передвигаться. Приходится сделать вывод, что рачки продолжают передвигаться, пока не найдут свободное место». Выяснилось также, что личинки S. balanoides распределяются равномерно по отношению к молоди и взрослым особям только своего вида и близких видов, но садятся в непосредственной близости или даже прямо на домик представителя другого семейства — Chthamalus stellatus. Это приводит к ожесточенной конкуренции между Chthamalus и Semibalanus.
Личинки некоторых видов оседают только на других животных. Есть виды, живущие исключительно в губках (Acasta spongicola, Balanus spongicola и др.) или главным образом на раковинах моллюсков и на панцирях ракообразных, в том числе камчатского краба (Solidobalanus hesperius в Тихом океане).
В процессе дальнейшего роста молодых морских желудей степень густоты их поселения очень важна, так как определяет форму их домика. В густых поселениях часто он приобретает «ли-лиевидную» форму, т. е. его диаметр в верхней части больше, чем в нижней, в разреженных поселениях он конический. Возможно, что на форму домика влияют и другие факторы.
Морские желуди растут довольно быстро. В тропиках некоторые виды достигают полово-зрелости через 8—16 суток после оседания. Обычному в Балтийском, Черном и Азовском морях В. improvisus, а также дальневосточному S. са-riosus для этого требуется 3 месяца. Продолжительность жизни разных видов колеблется от 1—2 до 5 — 7 и более лет.
Как уже указывалось, морские желуди и морские уточки населяют самые различные глубины. Некоторые виды хорошо приспособлены к существованию в приливо-отливной зоне. Они активны только во время прилива, а когда вода отступает, они плотно закрывают крышки домика и погружаются в состояние, сходное с анабиозом. При переносе в аквариум рачки в течение нескольких дней сохраняли тот же ритм, прекращая биение ножек в часы отлива. Однако, даже находясь вне воды, животные продолжают дышать, хотя и менее интенсивно, чем под водой. Только
при наступлении резко неолагоприятных условии они впадают в состояние «скрытой жизни» и в течение нескольких месяцев сохраняют жизнеспособность в сухом состоянии, без вреда для себя подвергаются длительному действию пресной воды, а в условиях опыта переносят заливание вазелиновым маслом, содержание в азоте и водороде, т. е. продолжают существовать при отсутствии кислорода.
Этой способностью к «скрытой жизни» объясняется, между прочим, появление морских желудей (В. improvisus и В. eburneus) в 1955 г. в Каспийском море, где их раньше не было. Они, несомненно, переселились туда из Черного и Азовского морей на днищах судов, прошедших по Волго-Донскому каналу, и сумели пережить длительное воздействие пресной воды. Вероятно, несколько легче было преодолеть еще более длительное расстояние австралийскому морскому желудю Eiminius modestus, появившемуся на побережье Великобритании только в 1943 —1944 гг. Он также проделал этот путь на днищах судов, но оставаясь все время в морской воде, а затем стал быстро расселяться вдоль берегов Европы, вытесняя местные виды — S. balanoides и В. improvisus. Однако при пониженной солености В. improvisus преобладает над новым вселенцем, а при низкой температуре преимущество получает S. balanoides.
Выше остальных морских желудей поселяются виды рода Chthamalus. В Черном море Ch. stel-latus (табл. 50, 10) часто встречается на прибрежных скалах, выше уреза воды, довольствуясь брызгами, которые иногда до него долетают. Другие морские желуди, в частности обычные виды рода Balanus, образуют массовые скопления в пределах приливо-отливной полосы и во время прилива оказываются под водой. Целый ряд видов морских желудей, как, например, крупный (до 20 см высотой) дальневосточный Chirona everman-ni (табл. 50, 8), и морских уточек приурочен к области материкового склона. На больших океанических глубинах преобладают морские уточки из семейства Scalpellidae. Наибольшая глубина нахождения 7420 — 7880 м (Arcoscalpellinae gen. sp.).
Некоторые морские уточки прикрепляются не к дну, а к плавающим в воде предметам — кускам дерева, судам, а также к различным животным. Таковы виды родов Lepas и Conchoderma. Обычный широко распространенный вид С. auritum встречается и на кусках дерева, и на днищах судов, и на китах, морских черепахах и медленно плавающих рыбах. При этом С. auritum всегда садится только на твердую поверхность. На китах эта морская уточка находится только на прикрепившихся к их коже морских желудях Сого-nula (табл. 50, 5) и на китовом усе, а у кашалотов — на зубах, образуя целые гроздья. Уховид-ные придатки рачка, служащие, вероятно, для
отвода профильтрованной воды, сильнее разв) у особей, сидящих на быстро движущихся жш ных.
Практическое значение The са определяется прежде всего тем, что морс: желуди составляют основу обрастания cyj Как уже указывалось, обросшее морскими: лудями (и другими животными) судно теряет ходную скорость. Соответственно увеличиваю время рейса, затраты на топливо и все осталы эксплуатационные расходы. Корабли перш чески приходится очищать от обрастания, а обходится очень дорого. Подсчитано, что то{ вый флот США ежегодно терпит убытки от обр тания, превосходящие 100 млн. долларов.
Во многих странах усиленно работают проблемой защиты судов от обрастания, в чг ности над изобретением таких красок для дв кораблей, которые были бы ядовиты для осед; щих на суда личинок морских желудей. Некс рые успехи в этом направлении достигнуты, «необрастающие» краски очень дороги.
Морские желуди вредят не только судам. П никнув в Каспийское море, они стали оседать ставных сетях, неводах и других рыболовных о днях и принесли немало неприятностей местн рыбакам, которые раньше с ними не стал! вались.
С другой стороны, несомненно и положителы значение морских желудей и морских уточек. личинки нередко составляют существенную ча| прибрежного морского планктона и в большом] личестве поедаются некоторыми планктонояд[ ми рыбами. Так, у берегов Норвегии весной к да питается преимущественно личинками ycoi гих, поедает их и беломорская сельдь, а так некоторые другие рыбы. Взрослые усоногие в| лой степени используются рыбами, которые ч то не могут оторвать их от субстрата и раскуси1 Тем не менее морские желуди отмечены в качеа объектов питания трески, пикши, у осетра и \
Человек также не брезгует усоногими. Из кр) ного морского желудя Megabalanus psittacus Чили делают консервы. В Испании и Порт)ч лии добывают морскую уточку Pollicipes pollil pes, которая используется и в пищу, и для прш товления тука. Многие другие усоногие тага несомненно, съедобны. Тур X е й е р д а л, ш сывая свое замечательное путешествие на «га Тики», рассказывает, между прочим, о том, к он и его спутники варили суп из морских ути которыми быстро обрастал их плот, и высоко oj нивает вкусовые качества этих рачков. Весы возможно, что в будущем человек станет исшш зовать усоногих в большей степени, чем ныне.
Большой урон наносят эти усоногие раки и обрастатели водоводов, подающих воду для охли дения различных агрегатов тепловых электроси ций, заводов и пр.
| РАКООБРАЗНЫЕ |
j ПОДОТРЯД АКРОТОРАЦИКОВЫЕ (ACROTHORACICA)
Занимаясь усоногими ракообразными, Ч. Дар-га открыл 'один сильно отличающийся по строению и образу жизни от морских желудей и мор-бких уточек вид, сверлящий раковины моллюс-ков, который он считал необходимым выделить i особый подотряд. В настоящее время таких видов известно более 40. Они делают ходы в рако-вннах брюхоногих, двустворчатых и панцирных шюсков, а также используют в качестве из-шкового субстрата скелет кораллов, мшанки, 1 иногда и домики морских желудей. Обычно Acrothoracica селятся в пустых раковинах уже погибших моллюсков, часто в раковинах, занятых рами-отшельниками, или в мертвом коралловом полшшяке, но известны и случаи поселения Bemdtia purpura в живых кораллах. Будучи хорошо защищены известковой толщей раковины моллюска или скелета коралла, они, естественно, лишены собственных известковых иастинок, свойственных их только что рассмотренным родичам. Только на передней части го-рвы самки есть хитиновая пластинка, принимаю-
вероятно, участие в процессе сверления.
г ротового отверстия расположены пара |шл и две пары челюстей, а позади рта пара передних грудных ножек (ротовые цирри). На значительном расстоянии от них находятся собранные |ьодну группу остальные 3—5 пар одноветвистых ш двуветвистых грудных ножек (терминальные ррри). Грудные ножки совершают около 60 взма-ювв 1 мин, но не высовываются при этом за края наши, как это бывает у морских желудей и юрских уточек. Тем не менее благодаря взмахам рудных ножек вода внутри мантийной полости тика все время сменяется и приносит достаточ-|ие количество мелких пищевых частиц. Они по-йупают в кишечник, либо сквозной, либо лишен-|и заднего отдела и ануса, но зато снабженный
'шогочисленными ветвящимися слепыми выроста-п, заходящими в голову, в грудные ножки и в fугие части тела. _ Acrothoracica — раздельнополые животные. Кар-инковые (0,4—2 мм) самцы живут поодиночке, ■крепившись к мантии самки (длина самок иногда достигает 20 мм). Они обычно сохраняют ■Ьик циприсовидной личинки или имеют очень ррощенное строение. У самцов отсутствует пи-■рварительный тракт, и, по-видимому, они сущест-■ротзасчет питательных веществ, сохраршвших-|и от личиночного периода. Развитие — с мета-И|озом, из яйца у большинства видов вылупля-1и циприсовидная личинка, а стадии науплиу-и и метанауплиуса протекают под защитой яй-■вых оболочек. Циприсовидная личинка садит-1ш подходящий субстрат, сбрасывает раковину ■ превращается в сформировавшегося рачка.
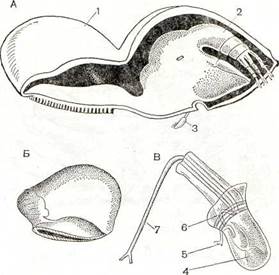
Рис. 223. Строение Tripetosa lampos:
А — взрослая самка с карликовым самцом; Б — молодая самка, передняя часть тела которой еще не превращена в диск; -В — самец при большом увеличении; 1 — передний конец тела; 2 — грудной отдел с ножками; 3 — самец; 4 — семенник; 5 — передняя антенна; б — лопасти мантии; 7 — пенис.
Рачок приступает к сверлению известняка, пользуясь хитиновой пластинкой и выделяя вещество, растворяющее соли кальция.
Acrothoracica широко распространены в Мировом океане. Ископаемые формы известны начиная с карбона.
ПОДОТРЯД КОРНЕГОЛОВЫЕ РАКООБРАЗНЫЕ (RHIZOCEPHALA)
Еще больше отличаются от обычных усоногих раков паразитические корнеголовые, организация которых предельно упрощена под воздействием паразитизма. Взрослые представители этого подотряда не сохраняют никаких признаков членистоногих. Они живут в десятиногих ракообразных — в крабах, раках-отшельниках и некоторых креветках.
Все тело краба, зараженного корнеголовым рачком саккулиной (Sacculina), пронизано ветвящимися, как корни растений, отростками паразита. Они оплетают все внутренние органы хозяина — кишечник, половые железы, нервную систему, внедряются в мускулатуру. Этими отростками рачки высасывают соки краба. Отростки впадают в обширный мешок, расположенный под брюхом хозяина на поверхности его тела. В мешке находятся сильно развитые яичники и маленькие семенники. Все остальные органы — кишечник, органы чувств и органы выделения — отсутствуют.
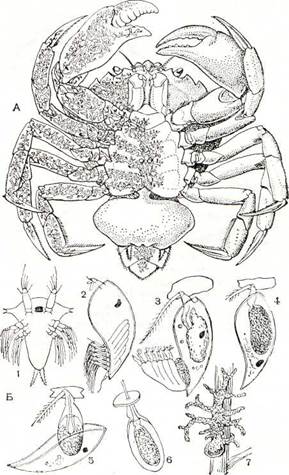
Рис. 224. Sacculina carcini.
Л — взрослый рачок, паразитирующий в крабе Carcinus maenas (левая половина тела краба изображена прозрачной). В — развитие личинки: 1 — науплиус; г — циприсовидная личинка; 3 — ее прикрепление и первая линька; 4 — строение личинки после линьки; 5 — вторая линька и формирование кентрогона; 6 — миграция клеток кентрогона в тело краба; 7 — молодой паразит на поверхности средней кишки краба.
Систематическое положение этого существа, так же как и в других подобных случаях (например, описанного выше дендрогастера), выясняется при изучении его развития (рис. 224, Б). Из яиц, откладываемых в полость мешка, выходит типичный для усоногих науплиус, выбирающийся наружу и плавающий в воде. Через несколько линек он превращается в крупную циприсовидную личинку. Личиночные стадии корнеголовых отличаются от соответствующих им стадий других усоногих главным образом отсутствием кишечника. Они питаются за счет содержащихся в их теле запасов желтка. Проплавав 3—4 дня, циприсовидная личинка прикрепляется передними антеннами к основанию щетинок конечностей или брюшной стороны тела молодых крабов. После этого она испытывает удивительные превращения. При очеред-
ной линьке она теряет большую часть своего те со всеми конечностями, но сохраняет маня раковину и глаз, а затем линяет еще раз и отб| сывает и их. От личинки остается малены овальное мешковидное тельце, внутри котор го располагаются недифференцированные клетн имевшиеся еще у науплиуса. Эта стадия пол чила название стадии кентрогона. Покр вы и передние антенны кентрогона образуют ш низанный каналом вырост, пробуравливающ покровы краба. Через этот канал кучка неди ренцированных клеток проникает в тело краба вероятно, кровью хозяина переносится на повел ность его средней кишки.
Здесь начинается рост паразита. Отростки а тела удлиняются, разветвляются и начинают i летать внутренние органы краба. Они не раз» тают ни кишечника, ни половых желез хеш на, всасывая питательные вещества из омыв» щей их крови и выделяя в организм хозяина! бросы своей жизнедеятельности. Через 7—81 сяцев паразит образует мешковидный вырог Когда он достигнет 2 мм в поперечнике, он п[ рывает покровы хозяина и выпячивается наруж а затем продолжает расти в течение 6 недел увеличиваясь до 12 мм в диаметре. К этому щ мени в мешковидном выросте заканчивается с] ревание половых желез, которые начинают фув ционировать. Кроме того, здесь же формируен очень слабо развитая нервная система. Всего ■ развития взрослой саккулины требуется, таи образом, 9 месяцев с момента прикрепления к■ зяину циприсовидной личинки. Паразит жш около 3 лет, и хозяин обычно переживает ег«продолжает существовать после смерти сам лины.
Присутствие саккулины оказывает мощное м действие на организм краба. Заражение сая линой молодого самца подавляет развитие половых желез. С каждой последующей линь» он становится все более похожим на самку: бри ной отдел его расширяется, исчезают преврая ные в совокупительный орган передние брюки конечности, меняется характер щетинок брюш Японские зоологи Ишикава и Янам м а ч и описали зараженного саккулиной cal краба Eriocheir japonicus, семенники котор«под влиянием паразита переродились в яичнн продуцировавшие нормальные яйца. Если 1 кулина заразила взрослого крупного самца i самку, это не приводит к изменению вторично! ловых признаков хозяина, но нарушает деята ность его половых желез — происходит пари тарная кастрация.
Нет сомнения в том, что причиной таких щ нений в организме краба служит воздейсти саккулины на его органы внутренней секрец! Непосредственно или выделяя специфические! щества, паразит подавляет выделение гормон!
 йивующих на формирование вторичпополовых рнаков самца и контролирующих деятель- ркть половых органов самца и самки.
йивующих на формирование вторичпополовых рнаков самца и контролирующих деятель- ркть половых органов самца и самки.
зиты раков-отшельников Peltogaster и stfrella также вызывают паразитарную каст- своих хозяев, но заражение ими самцов № приводит к развитию у них вторичнополовых лрюнаков самок. В отличие от саккулины у этих вразитов есть дополнительные самцы, сохра-шощие облик циприсовидных личинок. Семен-ики гермафродитных особей очень малы и не функционируют. Самцы прикрепляются перед-|1вш антеннами к мешковидной части гермафро-вгных особей, и масса заполняющих их недифференцированных клеток переселяется в недоразвитый семенник гермафродита. Часть этих клеток зреобразуется в сперматозоиды. При линьке раков-отшельников наружные мешковидные части их паразитов отваливаются вместе с покровами хозяина. Однако это не приводит I гибели паразита: из корневидной, находящейся внутри рака-отшелыгака части паразита вновь врастает его мешковидная часть. У Chologaster наблюдалось четырехкратное отпадение мешков рамена их новыми.
Способность к почкованию особенно ярко прошляется у Thompsonia — паразита креветок из иейства раков-щелкунов. На теле зараженной креветки всегда находится большое число мешко-цдных паразитов, объединенных общей корневой ■темой. Она возникает из одной личинки, а затем образует много мешковидных особей. Корнеголовые ракообразные широко распространены во всех морях и океанах, за исключением йодных южных и северных морей. Есть они и Черном море, и в наших дальневосточных морях. I) Северной Атлантике, Средиземном и Черном иорях около 6% представителей массовых видов крабов заражено саккулиной. В тропиках некоторые корнеголовые вместе со своими хозяевами «битают и в пресных водах. Всего известно около 250 видов этих удивительных ракообразных.
ПОДКЛАСС РАКУШКОВЫЕ
РАКООБРАЗНЫЕ
К этому подклассу принадлежат большей частью ркие (обычно мельче 1 мм, в редких случаях до 30 мм) рачки, обладающие двустворчатой, час-га пропитанной солями кальция раковиной, це-шком покрывающей их тело. Между створками раковины при движении животных высовываются галько антенны и иногда концы ножек, а также ■рка. Тело очень сильно укорочено и утратило
сегментацию. Число грудных • ножек сокращено до 1—3 пар. Все головные придатки и грудные ножки очень сильно специализированы: каждая пара выполняет какую-нибудь определеиную функцию. Сердца и жабр в подавляющем большинстве случаев нет. Из яйца выходит науплиус, имеющий, как и взрослые особи, двустворчатую раковину.
Ракушковые обитают как в море, так и в пресных водоемах. Большинство видов этих рачков передвигается по дну и водным растениям, но среди морских видов есть и планктонные. Благодаря прочной раковине ракушковые хорошо сохраняются в ископаемом состоянии и используются геологами для определения возраста различных отложений. Ракушковые — очень древняя группа животных: их остатки известны с нижнего кембрия.
Одни исследователи считают, что подкласс включает единственный отряд, имеющий то же название, другие разделяют его на 4 отряда. Всего известно более 2000 ныне живущих и 12 000 ископаемых видов.
Трудно найти водоем, где не жили бы ракушковые рачки. По разнообразию условий, к которым они приспособились, их можно сравнить только с веслоногими. В мелких лужах, в прудах, в крупных озерах, в подземных водах, на всех глубинах Мирового океана, как на дне, так и в толще воды можно обнаружить этих невзрачных мелких животных. Более того, известны 7 видов (Mesocyp-ris terrestris, Scotia audax и др.), которые перешли к сухопутному существованию: они живут в лесной подстилке влажных лесов Южной Африки, Мадагаскара, Австралии, Тасмании и Новой Зеландии. Как у многих других видов ракушковых, створки их раковин покрыты щетинками, что в данном случае способствует удержанию влаги и препятствует высыханию. Ракушковые из рода Potamocypris обнаружены в теплых водах ручья в штате Орегон в США при температуре 54 °С. Среди ракушковых имеются и комменсалы, живущие на разных животных, например на сверлящем дерево равноногом раке рода Limnoria.
Несмотря на столь разные условия существования и различный образ жизни (среди ракушковых есть хищники, есть и грунтоеды, донные и планктонные формы, зарывающиеся в грунт и ползающие по растениям и т. д.), организация ракушковых в общем довольно однообразна. Все они обладают двустворчатой раковиной, соответствующей кара-паксу других ракообразных. Внутренняя ее поверхность хитинизирована, а наружная обычно известковая. Левая и правая створки раковины соединены на спинной стороне эластической связкой. У многих видов есть и замок, состоящий из выступов па одной створке и соответствующих им углублений на другой. Приблизительно в середине тела расположен мощный мускул-замыка-
тель, прикрепленный к правой и левой створкам. При сокращении этого мускула раковина плотно закрывается.
Форма и строение раковины зависят от образа жизни. У Зрачков, живущих на поверхности жесткого грунта, раковина обычно толстостенная и снабжена различными буграми и выростами; у видов, роющихся в грунте, она толстостенная, но гладкая, а у видов, ползающих по растениям и живущих на мягком илистом грунте, она тонкая и гладкая. В особенности тонкой раковиной обладают морские планктонные виды, плотность тела которых приближается к плотности воды.
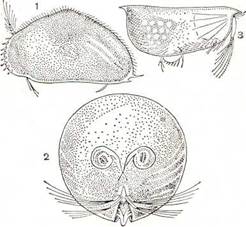
Голова часто хитинизирована. На ней обычно помещается непарный науплиальный глаз, а у представителей морского семейства Cypridinidae имеется пара сложных фасеточных глаз, достигающих у глубоководного планктонного Gigan-tocypris очень крупных размеров (рис. 225, 2). Участки раковины, находящиеся перед глазами, прозрачны, так что рачки видят через раковину.
На голове имеется четыре пары придатков. Наиболее мощная из них — задняя антенна, служащая основным органом передвижения. Она богата различными щетинками, приобретающими у некоторых видов форму маленьких клешней. У многих пресноводных ракушковых передняя антенна также длинная, хотя и тоньше задней. Они плавают, взмахивая передними антеннами вперед и вверх, а задними вперед и вниз, двигаясь при этом по равнодействующей, т. е. прямо. У представителей преимущественно морского семейства Cythe-ridae на конце редуцированной наружной ветви задних антенн открываются протоки парной паутинной железы. Вертикально опускаясь с какого-нибудь подводного предмета, рачки выделяют, как пауки, паутинную нить, а затем могут ею пользоваться при возвращении назад. Эта нить позволя-
Рис. 225. Ракушковые ракообразные:
1 — Cypris pubera; 2 — Gigantocypris; 3 — Conchoecia elegans.
ет им также удерживаться на грунте и так npoi востоять течению.
Жвалы снабжены сильно развитым щупике который нередко помогает движению рачи Некоторые Cypridmidae, например Asterope,; рываются в грунт при помощи щупиков яш Передние челюсти у разных групп ракушков; устроены различно. Часто их наружная вет превращена в широкую пластинку, взмахи ког рой сменяют воду в полости раковины. Дыхан осуществляется через стенку тела, и только Asterope на заднем отделе тела есть специальн выросты покровов, функционирующие как щ ры. У большинства ракушковых челюсти сыабл ны специализированными щетинками и участву] в захватывании пищи. Задние челюсти, по мнеш большинства исследователей, у ракушковых (сутствуют, но некоторые считают, что первая па грудных ног у них в действительности являет второй парой челюстей.
Первая, а у некоторых семейств (Cytherids Darwinulidae) и вторая пара грудных ног таю ходильные, но у Gypridae и у всех представи лей морского подотряда Myodocopa задние груда ножки загнуты на спинную сторону и приспосо лены для очистки мантийной полости: они ул ляют попавшие туда мелкие посторонние частя Есть роды, совсем лишенные обеих пар (Polycop или задней пары (Cytherella) грудных ножек. За няя часть тела в большинстве случаев закат вается хорошо развитой фуркой, принимают участие в движении рачка.
Эти движения очень разнообразны. Многие пре новодные Cypridae могут и плавать и ползат При ползании они цепляются щетинками задш антенн за субстрат и отталкиваются от него пере ними грудными ногами и фуркой; передние ai тенны при этом протянуты вперед и нащуньши путь (рис. 226, Б). Некоторые виды (Notodrom monacha, Heterocypris incongruens, Cyproidi marginata) могут передвигаться брюшной сторон! вверх, подвешиваясь снизу к поверхностной ша ке натяжения, как это делает описанный вышеш вистоусый рачок Scaphoieberis. Другие способ! зарываться в грунт на глубину до 7 см. Виды, щ способленные к передвижению по поверхнои грунта и растений, часто отличаются удлинени ми задними антеннами и передними грудными hoi ками. Морские планктонные Conchoecia и Gigaij tocypris плавают при помощи мощных задш антенн.
Также различны и способы питания. Мня ракушковые питаются преимущественно водор> лями, остатками растений и животных и оргаш ческим веществом грунта (Cytheridae, Darwintl dae, Candonidae, Cypridopsis). Среди моря видов есть хищники. В кишечнике Gypridii castanea обнаружены остатки мизид и киленоМ моллюсков, в кишечнике С. norvegica — остав
шихет. Планктонные ракушковые поедают других планктонных животных, в первую очередь клоногих, но иногда и щетинкочелюстных и ■иких рыб, а некоторые виды Gonchoecia наряду |(зоопланктоном используют и фитопланктон. Некоторые роды (морские Asterope, Cyclasterope, iCytJierella, пресноводный Notodromas) — настоя-Le фильтраторы с соответственно устроенными ротовыми придатками. Asterope и Cyclasterope врываются в грунт, выставляя над ним концы кредних антенн, образующие отверстие в грунте,, ррез которое проходит служащий для фильтра-|р ток воды.
Все ракушковые раздельнополы. Иногда на-июдается половой диморфизм: самки могут отжаться от самцов строением карапакса-рако-|иы, глаз или задних антенн. У многих прес-Ьводных Cypridae самки размножаются партено-тетически, а самцы вообще неизвестны. В ус-■овиях аквариума Herpetocypris reptans содер-шп в течение 30 лет, причем все это время про-(подило партеногенетическое размножение и сам-то ни разу не появились. Есть виды, которые i северной части области распространения раз-иожаются партеногенетически, а в южной встречаются и самцы и самки и наблюдается процесс водотворения.
Половые органы ракушковых устроены очень (сложно. Одной из самых удивительных особен-1юсгей этих рачков можно считать огромные размеры их сперматозоидов, превосходящих по вели-шк сперматозоиды всех остальных животных. Например, у ракушкового рачка Pontocypris ]иной 0,7 мм длина сперматозоида достигает 6 мм, т. е. оказывается в 8 раз больше самого животного. Длина сперматозоида человека 1(0,062 мм) почти в 100 раз меньше, чем длина спер-штозоида рачка.
I Вмещающие такие гигантские сперматозоиды ш.товые пути самцов и самок представляют собой [аяьно извитые каналы. Самцы имеют сложный шгулятивный аппарат, при помощи которого через щель раковины вводят сперматозоиды в по-ивое отверстие самки. Яйца оплодотворяются jipz выходе из полового отверстия и сначала ока-раются в полости раковины, в ее задней части. ■Через некоторое время самка откладывает яйца |и камни, поверхность грунта или на растения. Некоторые Cytheridae и Darwinulidae носят яй-ni под раковиной до выхода из них личинок, а у Itypridmidae под раковиной полностью формируются молодые рачки, после чего мать удаляет In наружу своими грудными ногами. I У остальных ракушковых из яйца выходит науп-«с своеобразного строения. Он снабжен двуст-риатой раковинкой, более короткой и сильнее су-шающейся к заднему концу, чем раковинка шослых рачков. Его задние антенны и жвалы рветвисты, причем последние заканчиваются
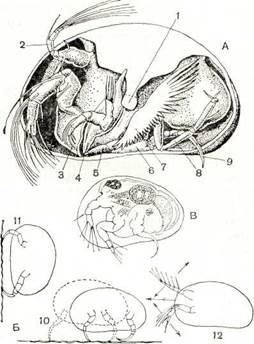
Рис. 226. Ракушковые ракообразные:
А — строение Cypridopsis vidua; Б — движение Cypris; В — на-уплиус; 1 — мускул-замыкатель раковины; 2 — передняя антенна; 3 — верхняя губа; 4 — жвала; б — передняя челюсть; 6 — задняя челюсть; 7 — наружная ветвь передней челюсти; 8 — задняя грудная нога; 9 — передняя грудная нога; 10 — ползание по дну; 11 — ползание по растениям; 12 — плавание.
длинным когтем. Науплиус может не только хорошо плавать, но и ползать при помощи задних антенн и жвал. Личинка линяет, постепенно приобретает недостающие конечности и превращается во взрослого рачка. У пресноводных видов продолжительность личиночного развития колеблется от 5 недель до 4 месяцев. В отличие от других ракообразных взрослые ракушковые не линяют. Длительность их жизни различна у разных видов. Пресноводные виды, науплиусы которых вылупляются из перезимовавших яиц, живут обычно не более 4 месяцев, виды семейства Candoni-dae — 9 и более месяцев, а морской Philomedes globosus — не менее 2Va лет.
ПОДКЛАСС ВЫСШИЕ
РАКООБРАЗНЫЕ
(MALAGOSTRACA)
Основные признаки видов подкласса — постоянное число грудных и брюшных сегментов, а также присутствие брюшных конечностей. Грудь состоит из 8, а брюшко из 6 или 7 сегментов и
заканчивается тельсоном. Нередко передний или несколько передних грудных сегментов срастаются с головой и их конечности превращаются в ного-челюсти. Иногда срастаются между собой или с тельсоном и б^рюшные сегменты. Мужские половые отверстия всегда открываются на последнем, а женские на шестом грудном сегменте. Желудок подразделен на жевательную и фильтрующую камеры. Всегда есть сильно развитая пищеварительная железа, сердце и кровеносные сосуды.
Название «высшие» не совсем точно. Если постоянство числа грудных и брюшных сегментов можно считать признаком более высокой организации, чем их непостоянство, присущее другим подклассам ракообразных, то присутствие брюшных конечностей, наоборот, говорит о меньшей специализации, чем их исчезновение, наблюдающееся у других подклассов. По всей вероятности, высшие ракообразные развивались независимо от других подклассов, каждый из которых сохранил те или иные примитивные признаки, унаследованные от общих предков.
Представители высших ракообразных, как правило, крупнее, чем рачки, относящиеся к другим подклассам. Они распространены чрезвычайно широко: населяют самые разнообразные водоемы и частично приспособились к обитанию на суше. Систематика подкласса очень сложна: он делится на надотряды, которые в свою очередь подразделяются на отряды. Таких надотрядов насчитывается пять.
НАДОТРЯД ФЙЛЛОКАРИДЫ (РНУЫЬОСАШВА)
Этот надотряд включает единственный очень бедный видами отряд, сохраняющий признаки примитивной организации.