 2015-08-13
2015-08-13 511
511НС—<СН,)*-5—CHi
кислоп (8 - 12}
чачяоп (8 - J2)
МрКН
J
НС—<CHs)i—СООН
с
г
НС—СИ,—«ЮН
«=о
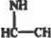
—он
| с=о |
| I НС—СН(СН,)—ОН с—о |
 трео*пвд
трео*пвд
| г ВС—СН,- и |
Гяупмин
| NH |
(5-8)
»исв, лейцв, ■эолсйшп,
| оон |
(40-5С)
Груши
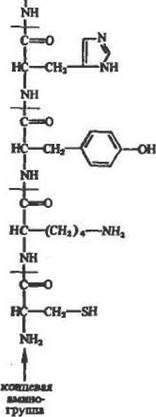
Рис. 13. Полипептидная цепь. Условно составлена из аминокислот в порядке
убывания числа химических реакций, в которых способны принимать участие
функциональные группы нх Соковых ряднкалоп, в скобках при названиях амнио-
кисло* указано среднее процентное содержание в белках
тате этого за счет гидрофобных взаимодействий полипептидные цепи сворачиваются в глобулярные структуры, ядро которых и составляют неполярные фрагменты полипептидных цепей, а наружный слой образуют звенья с полярными и ионогенными группами, такими, как —SH, —ОН, —СООН, —NH2. Конечно, некоторое количество гидрофобных группировок также может оказываться на поверхности; из л их на глобуле формируются контактные участки для связывания субстратов и {или) дли меж субъединичных взаимодействий. Ароматические аминокислотные остатки (тирозина' и триптофана) распределены между внутренними областями и наружным слоем так, чуо лишь часть из них оказывается доступной растворителю и, следовательно, растворенным в нем химическим реагентам.
Таким образом, с точки зрения возможностей иммобилизации ферментов белковые молекулы удобно представить в виде своеобразных шариков с экспонированными наружу функциональными группами: тнольными (цистенна), гидроксиль-ными (алифатическими — ссрина и треонина» ароматическими — тирозина)т карбоксильными (глутамнновон и аспарагнновой кислот» а также С-концевыми), гуанидиновыми (аргинина), имида-зольными (гистидина) и аминогруппами (е-лизина н а-т N- концевыми). Оценку количества тех или иных групп в различных белках в первом приближении можно сделать по данным, приведенным на рис. 13. Например, в таком небольшом белке, как трипсин или химотрипснн, с молекулярной массой около 25 000, должно быть до 10 остатков цистеина, около 30 алифатических ОН-груип, 3—7 остатков тирозина, 7—12 остатков аргинина, свыше 10 аминогрупп и 40 карбоксильных групп. Результаты такой оценки соответствуют известным экспериментальным данным.
Но, конечно» нет правил без исключений- В природе встречаются белки, вообще не содержащие некоторых из приведенных на рис. 13 аминокислотных остатков, например цистеина. В пепсине, протеолнтическом ферменте с молекулярной массой около 35 000 имеются не два десятка, как этого можно было бы ожидать, а всего лишь две аминогруппы: одна в-лизин а и одна tf-концевая. Но недостаток одних групп на поверхности белковой молекулы компенсируется другими. В общем, количество функциональных групп в белках, доступных модифицирующим агентам, достаточно велико и, казалось бы, нет особых проблем для ковалентной иммобилизации белковой молекулы с использованием хотя бы одной из этих групп. Тем не менее проблемы существуют и не малые. Дело в том, что в процессе ковалентной иммобилизации должны участвовать только те группы молекулы белка» которые не существенны для его функции (в нашем случае — катализа). С этой позиции попытаемся выявить в белке группы-мишени, наиболее предпочтительные для целей ковалентной иммобилизации. Для этого используем следующие критерии. Во-первыл, группы-мишени должны быть высокиреакциейнеспособными» чтобы по возможности обеспечить избирательность реакции модификации, а также ее протекание в мягких недена-турирующих условиях. Во-вторых, таких групп в белке должно быть достаточно много, чтобы обеспечить широкие возможности для Введения новых химических связей в белковую молекулу с регуляцией их числа и локализации и снизить таким образом вероятность модификации активных центров ферментов. Данные о сравнительной реакционной способности и относительному содержанию аминокислотных остатков приведены на рис. 13..
Самыми реакционноспособными группами в белках являются SN-группы цнстенна. Они способны принимать участие в разнообразных химических реакциях: окисления, ацилировання, алкилирования и т. д. Однако собственные тиольные группы бел-
ка не очень подходят на роль мишеней для кова лент ной иммобилизации. Дело не только в том, что содержание цистеина в белках в общем-то невелико. Очень часто в белках вообще нет сво: бодных SH -групп: они участвуют в образовании дисульфидных мостиков, стабилизирующих структуру ферментов. В тех же случаях, когда такие группы в белке есть, они, как правило, необходимы для каталитической активности. Тогда при иммобилизации возникает задача защитить SH-группы. Решить ее можно различными приемами, например обработкой белка я-оксимерку-рибензоатом: после окончания иммобилизации защитная группа отщепляется в слабокислой среде. Несмотря на указанные осложняющие обстоятельства использование SH-групп для кова-леитной иммобилизации ферментов остается привлекательным и в ряде случаев может быть осуществимо. Разработаны также методы, позволяющие вводить в молекулу белка экзогенные SH-группы, например, модификацией аминогрупп белка подходящими реагентами, в частности а цитированием тиолактоном гомо-цистеина.
Аминогруппы белка наиболее часто используют для различного рода химических модификаций и для целей ковалентной иммобилизации ферментов. Это обусловлено рядом причин. Во-первых, их и белке достаточно много (см. рис. 13). Во-вторых, аминогруппы высокореакционнеспособны и уступают лишь SH-группам по числу и разнообразию реакций, в которых они могут принимать участие. В-третьих, в большинстве своем аминогруппы играют второстепенную роль в поддержании структуры и функции ферментов. Важное свойство аминогрупп — способность прогонироваться (рК 9^—10), обеспечивает в физиологических условиях наличие на поверхности белков положительных зарядов, которые взаимодействуют с противоионами из раствора или с отрицательно заряженными карбоксильными группами белка, образуя в последнем случае солевые мостики. Если для нормального функционирования фермента важно сохранение положительного заряда, то этого можно достичь при химический модификации аминогрупп, переводя их (алкилированием) ил перыичиых во вторичные. В-четвиф-тых\ если некоторые аминогруппы, а не только их заряд, окажутся существенными для структуры а функции фермента, то их при необходимости нетрудно защитить, например, ацилированием ангидридом три фтору ксусной кислоты или м алей новым ангидридом (первая из указанных защитных групп снимается в щелочной среде, а вторая — в кислой). В-пятых, для ковалентной иммобилизации ферментов посредством их аминогрупп разработано большое количество подходящих носителей и сшивающих реагентов.
Таким образом, наиболее удобной группой — мишенью, следует считать аминогруппу. Однако, чтобы не сложилось впечатление об уникальности использования аминогрупп белков для целей ковалентной иммобилизации, необходимо отметить, что (как правило) обычные химические реагенты являются неспецифи
ческими и, помимо аминогрупп, в реакциях модификации могут принимать участие и другие функциональные группы белка, такие, как тиольные (цистеина), имидаэольные (гистидииа), гуаниднноаые (аргинина), гидроксильные (тирозина, серина и треонина), а также небелковые компоненты фермента или же реакционной системы, в частности вода. Реакционная способность тех или иных групп белка существенным образом зависит от условий: макро и микро. Группу макроусловий составляют такие, как природа растворителя» рН среды, температура —- эти условия могут и должны быть строго контролируемыми. Микроуслоикя, микросреда функциональной группы определяются структурой фермента, ее трудно изменять в нативиом белке, но важно учитывать при выборе метода и внешних условий модификации.
В заключение этого раздела необходимо подчеркнуть, что в целом белковые структуры весьма лабильны и существует огромное количество факторов, в том числе химических, вызывающих денатурацию и инактивацию ферментов. Поэтому исключительную значимость при операциях с белками приобретает выбор реакции и условий ее проведения: желательно, чтобы эти условия были мягкими и неденатурируюшими. Что же касается получения, трансформации и химической активации носителей и сшивающих агентов, то здесь практически нет ограничений в выборе условий реакции. Без особого преувеличения можно сказать, что любой твердый материал (природный или синтетический) может быть использован при необходимости в качестве носителя ковалентно иммобилизованных ферментов (см. гл. I).








