 2015-09-06
2015-09-06 11149
11149Окисление жирных кислот протекает в печени, почках, скелетных и сердечных мышцах, в жировой ткани.
Ф.Кнооп высказал предположение, что окисление молекулы жирной кислоты в тканях организма происходит в b-окислении. В результате от молекулы жирной кислоты отщепляются двууглеродные фрагменты со стороны карбоксильной группы. Процесс b-окисления жирных кислот складывается из следующих этапов:
Активация жирных кислот. Подобно первой стадии гликолиза сахаров перед b-окислением жирные кислоты подвергаются активации. Эта реакция протекает на наружной поверхности мембраны митохондрий при участии АТФ, коэнзима А (НS-КоА) и ионов Mg2+. Реакция катализируется ацил-КоА-синтетазой:
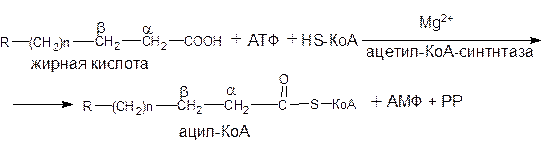
В результате реакции образуется ацил-КоА, являющийся активной формой жирной кислоты.
Транспорт жирных кислот внутрь митохондрий. Коэнзимная форма жирной кислоты, в равной мере как и свободные жирные кислоты, не обладает способностью проникать внутрь митохондрий, где, собственно, и протекает их окисление, переносчиком активированных жирных кислот через внутреннюю митохондриальную мембрану служит карнитин (g-триметиламино-b-оксибути-рат):
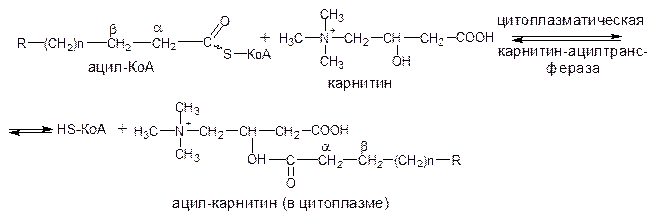
После прохождения ацилкарнитина через мембрану митохондрий происходит обратная реакция – расщепления ацилкарнитина при участии НS-КоА и митохондриальной карнитин-ацилтрансферазы:
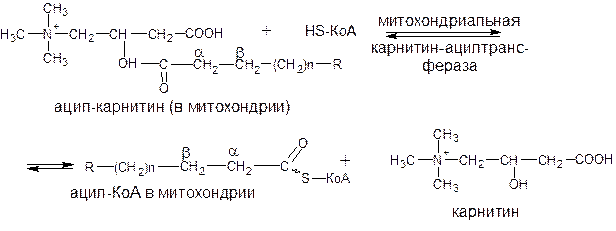
Ацил-КоА в митохондрии подвергается процессу b-окисления.
Этот путь окисления связан с присоединением атома кислорода к углеродному атому жирной кислоты, находящемуся в b-положении:
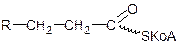
При b-окислении происходит последовательное отщепление от карбоксильного конца углеродной цепи жирной кислоты двууглеродных фрагментов в форме ацетила-КоА и соответствующее укорачивание цепи жирной кислоты:
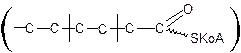
В матриксе митохондрии ацил-КоА распадается в результате повторяющейся последовательности четырех реакций (рис.8).
1) окисление с участием ацил-КоА-дегидрогеназы (ФАД-зависимой дегидрогеназы);
2) гидратация, катализируемой еноил-КоА-гидратазой;
3) второго окисления под действием 3-гидроксиацетил-КоА-дегидрогеназы (НАД-зависимой дегидрогеназы);
4) тиолиза с участием ацетил-КоА-ацилтрансферазы.
Совокупность этих четырех последовательностей реакций составляет один оборот b-окисления жирной кислоты (см. рис. 8).
Образовавшийся ацетил-КоА подвергается окислению в цикле Кребса, а ацетил-КоА, укоротившийся на два углеродных атома, снова многократно проходит весь путь b-окисления вплоть до образования бутирил-КоА (4-углеродное соединение), на последнем этапе b-окисления распадается на две молекулы ацетил-КоА.
При окислении жирной кислоты, содержащей n углеродных атомов, происходит n/2-1 цикл b-окисления (т.е. на один цикл меньше, чем n/2, так как при окислении бутирил-КоА сразу происходит образование двух молекул ацетил-КоА) и всего получится n/2 молекул ацетил-КоА.
Например при окислении пальмитиновой кислоты (С16) повторяется 16/2-1=7 циклов b-окисления и образуется 16/2=8 молекул ацетил-КоА.
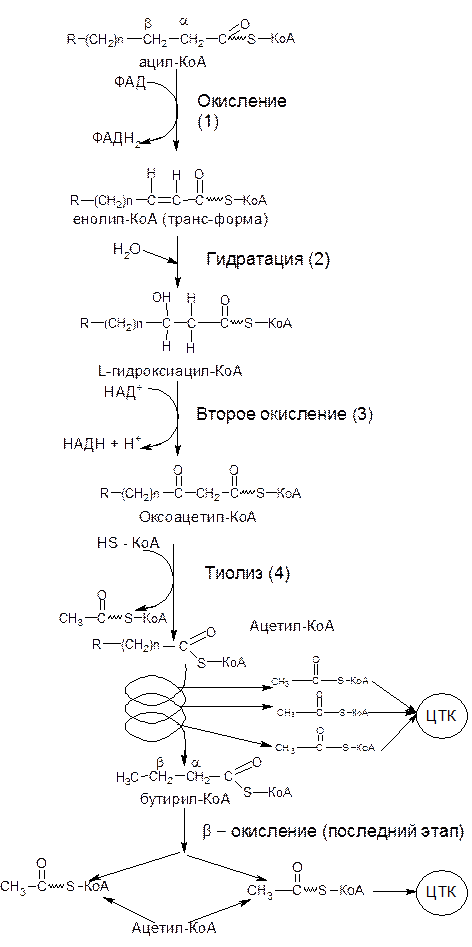
Рисунок 8 – Схема b-окисления жирной кислоты
Баланс энергии. При каждом цикле b-окисления образуется одна молекула ФАДН2 (см. рис. 8; реакция 1) и одна молекула НАДН+Н+(реакция 3). Последняя в процессе окисления дыхательной цепи и сопряженного с ним фосфорилирования дают: ФАДН2 – 2 молекулы АТФ и НАДН+Н+ – 3 молекулы АТФ, т.е. в сумме за один цикл образуется 5 молекул АТФ. При окислении пальмитиновой кислоты образуется 5*7=35 молекул АТФ. В процессе b-окисления пальмитиновой кислоты образуется 8 молекул ацетил-КоА, каждая из которых, «сгорая» в цикле Кребса, дает 12 молекул АТФ, а 8 молекул дадут 12*8=96 молекул АТФ.
Таким образом, всего при полном b-окислении пальмитиновой кислоты образуется 35+96=131 молекула АТФ. С учетом одной молекулы АТФ, потраченной в самом начале на стадии активации жирной кислоты, общий энергетический выход при полном окислении одной молекулы пальмитиновой кислоты составит 131-1=130 молекул АТФ.
Однако, образовавшийся в результате b-окисления жирных кислот ацетил-КоА, может не только окисляться до СО2, Н2О, АТФ, вступая в цикл Кребса, но использоваться на синтез холестерина, а также углеводов в глиоксилатном цикле.
Глиоксилатный путь специфичен только для растений и бактерий, у животных организмов он отсутствует. Данный процесс синтеза углеводов из жиров подробно описан в методическом указании «Взаимосвязь процессов обмена углеводов, жиров и белков» (см. п. 2.1.1, с. 26).

