 2017-11-30
2017-11-30 11752
11752Процесс реализации наследственной информации в биосинтезе осуществляется при участии трёх видов рибонуклеиновых кислот (РНК): информационной (матричной) – иРНК (мРНК), рибосомальной – рРНк и транспортной тРНК. Все рибонуклеиновые кислоты синтезируются на соответствующих участках молекулы ДНК. Они имеет значительно меньшие размеры, чем ДНК, и представляют собой одинарную цепь нуклеотидов. Нуклеотиды содержат остаток фосфорной кислоты (фосфат), пентозный сахар (рибозу) и одно из четырёх азотистых оснований – аденин, цитозин, гуанин, урацил. Азотистое основание – урацил - комплементарно аденину.
Процесс биосинтеза включает ряд этапов – транскрипцию, сплайсинг и трансляцию.
Первый этап называется транскрипцией. Транскрипция происходит в ядре клетки: на участке определённого гена молекулы ДНК синтезируется мРНк. В синтезе участвует комплекс ферментов, главным из которых является РНК-полимераза.
Синтез мРНК начинается с обнаружения РНК- полимеразой особого участка в молекуле ДНК, который указывает место начала транскрипции – промотора. После присоединения к промотору РНК-полимераза раскручивает прилежащий виток спирали ДНК. Две цепи ДНК в этом месте расходятся, и на одной из них идёт синтез мРНК. Сборка рибонуклеотидов в цепь происходит с соблюдением их комплементраности нуклеотидам ДНК, а также антипараллельно по отношению к матричной цепи ДНК. В связи с тем, что РНК-полимераза способна собирать полинуклеотид лишь от 5’-конца к 3’-концу, матрицей для транскрипции может служить только одна из двух цепей ДНК, а именно та, которая обращена к ферменту своим 3’-концом. Такую цепь называют кодогенной.
Антипараллельность соединения двух полинуклеотидных цепей в молекуле ДНК позволяет РНК-полимеразе правильно выбрать матрицу для синтеза мРНК.
Продвигаясь вдоль кодогенной цепи ДНК, РНК-полимераза осуществляет точное постепенное переписывании информации до тех пор, пока она не встречает специфическую нуклеотидную последовательность – терминатор транскрипции. В этом участке РНК-полимераза отделяется как от матрицы ДНК, так и вновь синтезированной мРНК. Фрагмент молекулы ДНК, включающий промотор, транскрибируемую последовательность и терминатор, образует единицу транскрипции – транскриптон.
Дальнейшие исследования показали, что в процессе транскрипции синтезируется так называемая про-мРНК – предшественник зрелой мРНК, участвующей в трансляции. Про-мРНК имеет значительно большие размеры и содержит фрагменты, не кодирующие синтез соответствующей полипептидной цепи. В ДНК наряду с участками, кодирующими рРНК, тРНК и полипептиды, имеются фрагменты, не содержащие генетической информации. Они получили название интронов в отличие от кодирующих фрагментов, которые называются экзонами. Интроны обнаружены на многих участках молекул ДНК. Например, в одном гене –участке ДНК, кодирующем овальбумин курицы, содержится 7 интронов, в гене сывароточного альбумина крысы – 13 интронов. Длина интрона бывает различной – от 200 до 1000 пар нуклеотидов ДНК. Интроны считываются (транскрибируются) одновременно с экзонами, поэтому пор-мРНК значительно длиннее, чем зрелая мРНК. Созревание, или процессинг, мРНК предполагает модифицирование первичного транскрипта и удаление из него некодирующих интронных участков с последующим соединением кодирующих последовательностей – экзонов. В ходе процессинга из про-мРНК специальными ферментами «вырезаются» интроны, а фрагменты экзона «сращиваются» между собой в строгом порядке. В процессе сплайсинга образуется зрелая мРНК, которая содержит ту информацию, которая необходима для синтеза соответствующего полипептида, то есть информативную часть структурного гена.
Значение и функции интронов до сих пор ещё не совсем выяснены, но установлено, что если в ДНК считываются только участки экзонов, зрелая мРНК не образуется. Процесс сплайсинга изучен на примере работы овальбумина. Он содержит один экзон и 7 интронов. Сначала на ДНК синтезируется про-мРНК, содержащая 7700 нуклеотидов. Затем про-мРНК число нуклеотидов уменьшается до 6800, затем – до 5600, 4850, 3800, 3400 и т.д. до 1372 нуклеотидов, соответствующих экзону. Содержащая 1372 нуклеотида мРНК выходит из ядра в цитоплазму, попадает на рибосому и синтезирует соответствующий полипетид.
Следующий этап биосинтеза – трансляция – происходит в цитоплазме на рибосомах при участи тРНК.
Транспортные РНК синтезируются в ядре, но функционируют в свободном состоянии в цитоплазме клетки. Одна молекула тРНК содержит 75-95 нуклеотидов и имеет довольно сложную структуру, напоминающую лист клевера. В ней выделяют четыре части, которые имеют особо важное значение. Акцепторный «стебель» образуется путём комплементарного соединения двух концевых частей тРНК. Он состоит из 7 пар оснований. 3’-конец это стебля несколько длиннее и формирует одноцепочечный участок, который заканчивается последовательностью ЦЦА со свободной ОН группой – акцепторный конец. К этому концу присоединяется транспортируемая аминокислота. Остальные три ветви представляют собой комплементарно спаренные последовательности нуклеотидов, которые заканчиваются неспаренными участками, образующими петли. Средняя из этих ветвей – антикодоновая – состоит из 5 пар и содержит в центре своей петли антикодон. Антикодон – это 3 нуклеотида, комплементарные кодону мРНК, который шифрует аминокислоту, транспортируемую данной тРНК к месту синтеза пептида.
Между акцепторной и антикодоновой ветвями располагаются две боковые ветви. В своих петлях они содержат модифицированные основания - дигидроуридин (D-петля) и триплет Т ᴪС, где ᴪ - псевдоуридин (Т ᴪС-петля). Между антикодоновой и Т ᴪС-ветвями содержится дополнительная петля, включающая от 3-5 до 13-21 нуклеотидов.
Присоединению аминокислоты к тРНК предшествует её активация ферментом аминоацил-тРНК-синтетазой. Этот фермент специфичен для каждой аминокислоты. Активированная аминокислота прикрепляется к соответствующей тРНК и доставляется ею на рибосому.
Центральное место в трансляции принадлежит рибосомам – рибонуклеопротеиновым органоидам цитоплазмы, во множестве в ней присутствующим. Размеры рибосом у прокариот в среднем 30*30*20 нм, у эукариот – 40*40*20 нм. Обычно их размеры определяют в единицах седиментации (S) – скорости осаждения при центрифугировании в соответствующей среде. У бактерий кишечной палочки рибосома имеет величину 70S и состоит из 2 субчастиц, одна из которых имеет константу 30S, вторая 50S, и содержит 64% рибосомальной РНК и 36% белка.
Молекула мРНК выходит из ядра в цитоплазму и прикрепляется к малой субчастице рибосомы. Трансляция начинается с так называемого стартового кодона (инициатора синтеза) – АУГ -. Когда тРНК доставляет к рибосоме активированную аминокислоту, её антикодон соединяется водородными связями с нуклеотидами комплементарного кодона мРНК. Акцепторный конец тРНК с соответствующей аминокислотой прикрепляется к поверхности большой субчастицы рибосомы. После первой аминокислоты другая тРНК доставляет следующую аминокислоту, и таким образом на рибосоме синтезируется полипептидная цепь. Молекула мРНК обычно работает сразу на нескольких (5-20) рибосомах, соединённых в полисомы. Начало синтеза полипептидной цепи называется инициацией, её рост – элогацией. Последовательность аминокислот в полипептидной цепи определяется последовательностью кодонов в мРНК. Синтез полипептидной цепи прекращается, когда на мРНК появляется один из кодонов – терминаторов – УАА -, - УАГ – или – УГА -. Окончание синтеза данной полипептидной цепи называется терминацией.
Установлено, что в клетках животных полипептидная цепь за одну секунду удлиняется на 7 аминокислот, а мРНК продвигается на рибосоме на 21 нуклеотид. У бактерий этот процесс протекает в 2-3 раза быстрее.
Следовательно, синтез первичной структуры белковой молекулы – полипептидной цепи – происходит на рибосоме в соответствии с порядком чередования нуклеотидов в матричной рибонуклеиновой кислоте – мРНК.
Биосинтез белка (трансляция) – важнейший этап реализации генетической программы клеток, в процессе которого информация, закодированная в первичной структуре нуклеиновых кислот, переводится в аминокислотную последовательность синтезируемых белков. Иными словами, трансляция – это перевод четырех буквенного (по числу нуклеотидов) «языка» нуклеиновых кислот на двадцатибуквенный (по числу протеиногенных аминокислот) «язык» белков. Перевод осуществялется в соответствии с правилами генетического кода.
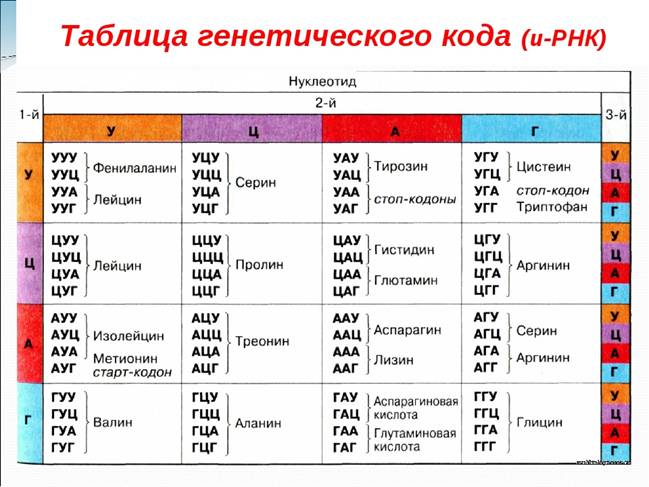
Важное значение для раскрытия генетического кода имели исследования М. Ниренберга и Дж. Маттеи, а затем С. Очоа и Г. Кораны, начатые ими в 1961г. в США. Они разработали метод и экспериментально установили последовательность нуклеотидов в кодонах мРНК, контролирующих местоположение данной аминокислоты в полипептидной цепи. В бесклеточную среду, содержащую все аминокислоты, рибосомы, тРНК, АТФ и ферменты, М. Ниренберг и Дж. Маттеи вводили искусственно синтезированный биополимер типа мРНК, представляющий собой цепочку одинаковых нуклеотидов – УУУ –УУУ –УУУ – УУУ - и т.д. биополимер кодировал синтез полипептидной цепи, содержащей только одну аминокислоту – фенилаланин; такая цепь называется полифенилаланином. Если мРНК состояла из кодонов, содержащих нуклеотиды с азотистым основанием цитозин – ЦЦЦ – ЦЦЦ – ЦЦЦ – ЦЦЦ -, то синтезировась полипептидная цепь, содержащая аминокислоту пролин, - полипролин. Искусственные биополимеры мРНК, содержащие кодоны – АГУ – АГУ – АГУ – АГУ -, синтезировали полипептидную цепь из аминокислоты серин – полисерин и т.д.
Обратная транскрипция — это процесс образования двуцепочечной ДНК на матрице одноцепочечной РНК. Данный процесс называется обратной транскрипцией, так как передача генетической информации при этом происходит в «обратном», относительно транскрипции, направлении.
Обратная транскриптаза (ревертаза или РНК-зависимая ДНК-полимераза) — фермент, катализирующий синтез ДНК на матрице РНК в процессе, называемом обратной транскрипцией.Обратная транскрипция необходима, в частности, для осуществления жизненного цикла ретровирусов, например, вирусов иммунодефицита человека и T-клеточной лимфомы человека типов 1 и 2. После попадания вирусной РНК в клетку обратная транскриптаза, содержащаяся в вирусных частицах, синтезирует комплементарную ей ДНК, а затем на этой цепи ДНК, как на матрице, достраивает вторую цепь.ретровирусы - это РНК-содержащие вирусы, в жизненный цикл которых входит стадия образования ДНК обратной транскриптазой и внедрение ее в геном клетки хозяина в форме провируса.
Предпочтительного места внедрения провируса в геном нет. Это позволяет отнести его к мобильным генетическим элементам.В состав ретровируса входит две идентичные молекулы РНК. На 5'-конце имеется Сap, на 3'-конце - поли А-хвост. Фермент обратную транскриптазу вирус "носит" c собой.
Геном ретровируса содержит 4 гена:gag-белок нуклеоида,pol-обратная транскриптаза,env-белок капсида (оболочки),онкоген.str5 = str3-короткий концевой повтор;U5, U3-уникальные последовательности,PB (primer binding site) - участок связывания затравки.На РВ садится (за счет комплементарности) tРНК и служит затравкой для синтеза ДНК.Синтезируется небольшой кусок ДНК.
Обратная транскриптаза, обладая еще и активностью РНК-азы Н, удаляет РНК в гибриде с ДНК, а за счет идентичности str3 и str5 этот одноцепочечный участок ДНК взаимодействует с 3'-концом второй молекулы РНК, которая служит матрицей для продолжения синтеза цепи ДНК.
Затем РНК-матрица уничтожается и по образовавшейся цепи ДНК строится комплементарная.
Образованная молекула ДНК длиннее РНК. Она содержит LTR (U3 str 3(5) U5). В форме провируса она находится в геноме клетки хозяина. При митозе и мейозе передается дочерним клеткам и потомкам.
Некоторые вирусы (такие как ВИЧ, вызывающий СПИД), имеют возможность транскрибировать РНК в ДНК. ВИЧ имеет РНК-геном, который встраивается в ДНК. В результате, ДНК вируса может быть объединено с геномом клетки-хозяина. Главный фермент, ответственный за синтез ДНК из РНК, называется ревертазой. Одной из функций ревертазы является создание комплементарной ДНК (кДНК) из вирусного генома. Ассоциированый фермент рибонуклеаза H расщепляет РНК, а ревертаза синтезирует кДНК из двойной спирали ДНК. кДНК интегрируется в геном клетки-хозяина с помощью интегразы. Результатом является синтез вирусных протеинов клеткой-хозяином, которые образуют новые вирусы

