 2018-01-08
2018-01-08 1077
1077Необходимые растениям макроэлементы и их физиологическая роль.
В растительном организме все процессы тесно взаимосвязаны. Исключение из питательной среды какого-либо необходимого элемента быстро вызывает изменение во многих, если не во всех, процессах метаболизма. В связи с этим выделить первичный эффект бывает чрезвычайно трудно. Сказанное относится в первую очередь к тем питательным элементам, которые не входят в состав определенных органических веществ, а играют скорее регуляторную или какую-то иную роль. В общем виде можно сказать, что питательные элементы имеют следующее значение: 1) входят в состав биологически важных органических веществ; 2) участвуют в создании определенной ионной концентрации, стабилизации макромолекул и коллоидных частиц (электрохимическая роль); 3) участвуют в каталитических реакциях, входя в состав или активируя отдельные ферменты. Во многих случаях один и тот же элемент может играть разную роль. Некоторые элементы выполняют все три функции.
Фосфор. Содержание фосфора в растениях составляет около 0,2% на сухую массу. Фосфор поступает в корневую систему и функционирует в растении в виде окисленных соединений, главным образом остатков ортофосфорной кислоты (Н2Р04-, HP042-, Р043-). Физиологическое значение фосфора определяется тем, что он входит в состав ряда органических соединений, таких, как нуклеиновые кислоты (ДНК и РНК), нуклеотиды (АТФ, НАД, НАДФ), нуклеопротеиды, витамины и многих других, играющих центральную роль в обмене веществ. Фосфолипиды являются компонентами биологических мембран, причем именно присутствие фосфата в их структуре обеспечивает гидрофильность, остальная часть молекулы липофильна. Многие витамины и их производные, содержащие фосфор, являются коферментами и принимают непосредственное участие в каталитических реакциях, ускоряющих течение важнейших процессов обмена (фотосинтез, дыхание и др.). Фосфор содержится в составе такого органического соединения как фитин (Са—Mg соль инозитфосфорной кислоты), который является основной запасной формой фосфора в растении. Особенно много фитина в семенах (до 1—2 % сухой массы). При всех превращениях в растительном организме фосфор сохраняет степень окисленности. На самом деле, все превращения сводятся лишь к присоединению или переносу остатка фосфорной кислоты (фосфорилирование и трансфосфорилирование). Фосфорилирование — это присоединение остатка фосфорной кислоты к органическому соединению с образованием эфирной связи, например взаимодействие фосфорной кислоты с карбонильной, карбоксильной или спиртовой группировками. Фосфорилирование белков осуществляется ферментами протеинкиназами и контролирует протекание обменных реакций в организме, включая синтез белка и РНК, регуляцию активности ферментов, и лежит в основе работы сигнальных цепей. Фосфорилироваться могут и другие соединения. Например, при фосфорилировании Сахаров образуются сахарофосфаты — эфиры Сахаров и фосфорной кислоты. Эти соединения, более лабильные и реакционноспособные, чем свободные сахара, играют существенную роль при дыхании, взаимных превращениях углеводов, их синтезе. Трансфосфорилирование — это процесс, при котором остаток фосфорной кислоты, включенный в состав одного органического вещества, переносится на другое органическое вещество. Ряд важнейших в биологическом отношении фосфорных соединений содержит несколько остатков фосфорной кислоты. Для фосфора характерна способность к образованию связей с высоким энергетическим потенциалом (макроэргические связи). Такие связи нестабильны, это облегчает их обмен и позволяет использовать энергию на самые различные биохимические и физиологические процессы. Важным соединением, содержащим макроэргические фосфорные связи, является АТФ. Фосфорная кислота, поступая в живые клетки корня, быстро включается в состав нуклеотидов, образуя АМФ и АДФ. Далее в процессе субстратного и окислительного фосфорилирования (анаэробная и аэробная фазы дыхания) образуется АТФ. По данным А.Л. Курсанова, уже через 30 с поступивший меченый фосфор (32Р) обнаруживается в АТФ. Образовавшаяся АТФ используется на активацию Сахаров, аминокислот, синтез нуклеиновых кислот, белков и на другие процессы. Недостаток фосфора влияет практически на все процессы жизнедеятельности растений. Для нормального протекания фотосинтеза, дыхания, роста требуется фосфор. В почве фосфор находится в малорастворимой форме, поэтому в обеспечении питания фосфором велика роль метаболизма корней. Поглощению фосфора способствует выделение корнями кислот, ферментов, углеводистых веществ.
Сера содержится в растениях в количестве 0,17%. Однако в растениях семейства крестоцветных ее содержание гораздо выше. Поступает сера в растения в виде сульфатиона S042-. Сера входит в состав органических соединений, играющих важную роль в обмене веществ организма. Так, сера входит в состав трех аминокислот — цистина, цистеина и метионина. Почти все белки включают аминокислоты, содержащие серу, поэтому становится понятна роль серы в белковом обмене организма. Сера входит также в состав многих витаминов и многих коферментов, таких, как биотин, тиамин, коэнзим А, глютатион, липоевая кислота и др. В связи с этим сера принимает участие в многочисленных реакциях обмена (аэробная фаза дыхания, синтез жиров и др.). В составе коэнзима А (СоА—SH) сера участвует в образовании макроэргической связи с ацильными группами кислот. Ацетилкоэнзим А (CH3CO~SCoA) играет роль в метаболизме углеводов, жирных кислот, аминокислот. Аденозилметионин используется при синтезе фитогормона этилена и лигнина. Сульфгидрильные группировки (SH) и дисульфидные связи (S—S) играют большую роль, обеспечивая взаимодействие между ферментами и их простетическими группами, а также участвуя в создании определенной конфигурации белковых молекул. Так, SH-группы связывают белок с такими коферментами, как НАД или ФАД. Часто за счет дисульфидных связей сохраняется трехмерная структура белка, а следовательно, его активность. Соединения серы участвуют в поддержании уровня окислительно-восстановительного потенциала клетки. Это относится к системам цистеин — цистин и SH—глутагион <-> S—S — глутатион. В составе белка тиоредоксина сера участвует в регуляции работы таких ферментов как Rubisco, АТФ-синтаза и др.. Сера входит в состав чесночных и горчичных масел. Именно с этим связан своеобразный вкус и запах некоторых растений семейства крестоцветные. Нельзя не отметить, что соединения серы, такие, как S-аденозилметионин, участвуют в образовании полиаминов, в частности спермедина. Согласно современным представлениям, полиамины играют большую и разностороннюю роль в жизнедеятельности организмов. Полиамины благодаря наличию заряженных аминогрупп во многих реакциях могут заменять неорганические катионы. Сера, поступая в растение в виде иона S042-, быстро переходит в органическую форму при участии АТФ и магния:
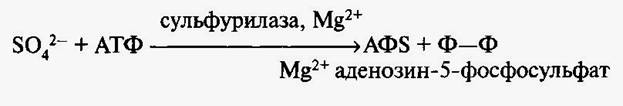
Такой активированный сульфат является короткоживущим соединением и через ряд этапов восстанавливается до цистеина при участии ферредоксина. Ферменты, участвующие в образовании цистеина, в частности сульфурилаза локализованы в цитозоле, пластидах и митохондриях. Из цистеина образуются цистин и метионин. Производным цистеина является глютатион, участвующий в передвижении серы по растению. Глютатион является сигналом для поступления иона S042- через корни. В восстановленной форме сера включается в аминокислоты. Восстановленная сера в растении может подвергаться снова окислению. Окисленная форма S042- неактивна. Показано, что в молодых органах сера находится главным образом в восстановленной форме, а старых — в окисленной. К числу необходимых растению металлов относят как макроэлементы К, Са, Mg, Fe, так и микроэлементы Си, Zn, Мл и др. Участие в каталитических реакциях характерно, главным образом, для металлов. Металлы могут осуществлять влияние на процессы обмена различным путем:
1) непосредственно входя в активный центр фермента (в простетическую группу или в апофермент). Таковы ферменты, содержащие железо, медь и некоторые другие элементы. Функция металла заключается чаще всего в переходе из восстановленной в окисленную форму и обратно, что сопровождается переносом электрона, например: Fe2+ —» Fe3+ + е;
2) активируя тот или иной фермент путем изменения заряда белка-фермента или его конфигурации;
3) являясь связующим мостиком между ферментом и субстратом и тем самым облегчая их взаимодействие;
4) изменяя константу равновесия ферментативных реакций;
5) изменяя равновесие между активной и неактивной формами фермента;
6) связывая ингибиторы тех или иных ферментативных реакций.
Кальций входит в состав растений в количестве 0,2%. В старых листьях его содержание доходит до 1 %. Поступает в виде иона Са2+. Роль кальция разнообразна. Кальций, соединяясь с пектиновыми веществами, дает пектаты кальция, которые являются важнейшей составной частью клеточных оболочек растений. Срединные пластинки, склеивающие клеточные оболочки соседних клеток, состоят по преимуществу из пектатов кальция. При недостатке кальция клеточные оболочки ослизняются, что особенно ярко проявляется в клетках корня. Кальций плохо передвигается по растению, поэтому для предупреждения ослизнения необходимо, чтобы ионы Са2+ непосредственно соприкасались с клетками корня. Сказанное было продемонстрировано в опытах, поставленных по методу изолированных водных культур. В этих опытах одну прядь корней помещали в питательный раствор, содержащий все необходимые питательные вещества; другую прядь корня того же растения — в раствор с исключением кальция.
Очень скоро клетки корня, которые находились в растворе без кальция, начали ослизняться и загнивать. Кальций повышает вязкость цитоплазмы, что видно на опытах с формами плазмолиза. В солях кальция плазмолиз имеет вогнутую форму, так как более вязкая цитоплазма с трудом отстает от клеточных оболочек. Присутствие кальция важно для нормального функционирования мембран. Дефицит кальция приводит к увеличению проницаемости мембран, нарушению их целостности, а соответственно процессов мембранного транспорта. Кальций принимает участие в поддержании структуры хромосом, являясь связующим звеном между ДНК и белком. При недостатке кальция наблюдаются повреждения хромосом и нарушение митотического цикла. Кальций необходим также для поддержания структуры митохондрий и рибосом, образования ламелл во вновь образующихся клетках. Кальций является активатором таких ферментов, как фосфорилаза, аденозинтрифосфатаза, дегидрогеназы, амилазы и др. Са2+ служит посредником для реакций растений на внешние и гормональные сигналы, входя в состав сигнальных систем. В этой связи большое значение имеет связывание Са2+ с белком кальмодулином, находящимся в цитозоле. В цитоплазме в обычных условиях поддерживается низкая концентрация кальция. При повышении внутриклеточной концентрации кальция в ответ на сигналы (внутренние и внешние) происходит его связывание с кальмодулином. Кальмодулин регулирует концентрацию Са2+ в клетке по принципу обратной связи. Комплекс Са2+ — кальмодулин способен влиять на активность ферментов, участвующих в синтезе и распаде циклических нуклеотидов (аденилатциклаза, фосфодиэстеразы), Са2+-зависимых протеинкиназ, Са2+-АТФазы. Кальмодулин влияет на активность структурных белков цитоскелета и таким образом контролирует перемещение органелл внутри клетки, изменение формы клетки, образование веретена деления. Кальций участвует в образовании клеточной стенки и росте растяжением. Кальций реагирует с различными органическими кислотами, давая соли, и тем самым является в определенной мере регулятором значения рН клеточного сока. Нейтрализуя щавелевую кислоту, образует характерные кристаллы щавелевокислого кальция.
Магний. Содержание магния в растениях составляет в среднем 0,17%. Магний поступает в растение в виде иона Mg2+. Магний входит в состав основного пигмента зеленых листьев — хлорофилла. Магний поддерживает структуру рибосом, связывая РНК и белок. Большая и малая субъединицы рибосом ассоциируют вместе лишь в присутствии магния, который также необходим для формирования полисом и активации аминокислот. Поэтому синтез белка не идет при недостатке магния, а тем более в его отсутствие. Магний является активатором многих ферментов. Важной особенностью магния является то, что он связывает фермент с субстратом по типу хелатной связи (клешневидная связь между органическим веществом и катионом). Так, например, присоединяясь к пирофосфатной группе, магний связывает АТФ с соответствующими ферментами. В связи с этим все реакции, включающие перенос фосфатной группы (большинство реакций синтеза, а также многие реакции энергетического обмена), требуют присутствия магния. Магний активирует такие ферменты, как ДНК- и РНК-полимеразы, аденозинтрифосфатазу, глютаматсинтетазу; ферменты, катализирующие перенос карбоксильной группы,— реакции карбоксилирования и декарбоксилирования; ферменты гликолиза и цикла Кребса, молочнокислого и спиртового брожений. В ряде случаев влияние магния на работу ферментов определяется тем, что он реагирует с продуктами реакции, сдвигая равновесие в сторону их образования. Магний может также инактивировать ряд ингибиторов ферментативных реакций.
Калий. Содержание калия в растении в среднем составляет 0,9%. Он поступает в растение в виде иона К+. Физиологическую роль калия нельзя считать полностью выясненной. Калий не входит ни в одно органическое соединение. Большая часть его (70%) в клетке находится в свободной ионной форме и легко извлекается холодной водой, остальные 30% в адсорбированном состоянии. В противоположность кальцию калий снижает вязкость протоплазмы, повышает ее оводненность, увеличивая гидратацию белков. Эта особенность действия калия хорошо проявляется в том, что в его солях плазмолиз имеет выпуклую форму, протоплазма легко отстает от клеточной оболочки. Следовательно, калий является антагонистом кальция. Соли калия растворимы и участвуют в регуляции осмотического потенциала клетки. В частности, большое значение имеет К+ в регуляции работы устьиц. Показано, что открытие устьиц на свету связано с накоплением в замыкающих клетках ионов калия. При этом К+ в обмен на Н+ поступает из клеток, окружающих устьичные. Корневое давление также во многом зависит от присутствия К+ в пасоке.
Калий активирует работу многих ферментных систем, например фермент, катализирующий фосфорилирование Сахаров, — гексокиназа, ферменты, катализирующие перенос фосфорной кислоты с пирувата на АДФ (пируваткиназа), а также ферменты, участвующие в образовании АТФ в процессе окислительного фосфорилирования. В связи с этим, по данным Э.И. Выскребенцевой, при недостатке калия резко падает содержание макроэргических фосфатов. Калий активирует и ряд ферментов цикла Кребса. Многие ферменты, участвующие в синтезе белка, требуют для своего действия присутствия калия. Несмотря на необходимость калия для проявления активности ряда ферментов, механизм этой активации до настоящего времени не представляется ясным. Недостаток калия замедляет транспорт сахарозы по флоэме. Влияние калия на передвижение органических веществ, по гипотезе Спаннера, проявляется благодаря образованию градиента электрического потенциала на ситовидных пластинках, который возникает при циркуляции калия между ситовидной трубкой и сопровождающими клетками. Влияние К+ на транспорт углеводов определяет его роль в формировании урожая. Под влиянием калия увеличивается накопление крахмала, сахарозы, моносахаридов.
Железо входит в состав растения в количестве 0,08%. Необходимость железа была показана в тот же период, что и остальных макроэлементов. Поэтому, несмотря на ничтожное содержание, его роль рассматривается вместе с макроэлементами. Железо поступает в растение в виде Fe3+, а транспортируется в листья по ксилеме в виде цитрата железа (III). Роль железа в большинстве случаев связана с его способностью переходить из окисленной формы (Fe3+) в восстановленную (Fe2+) и обратно. Железо входит в состав каталитических центров многих окислительно-восстановительных ферментов. В виде геминовой группировки оно входит в состав таких ферментов, какцитохромы, цитохромоксидаза, нитратредуктаза, нитритредуктаза, леггемоглобин, каталаза и пероксидаза. Цитохромная система является необходимым компонентом дыхательной и фотосинтетической электронтранспортной цепи. В силу этого при недостатке железа тормозятся оба этих важнейших процесса. Кроме того, целый ряд ферментов содержит железо в негемовой форме. К таким ферментам относятся некоторые флавопротеиды, нитрогеназа, железосодержащий белок ферредоксин, фитоферритин и др. Фитоферритин — является металлопротеидом, в виде которого железо аккумулируется в клетке. Железо необходимо для образования хлорофилла. При этом железо катализирует образование предшественников хлорофилла 5-аминолевулиновой кислоты и протопорфиринов. Предполагают, что железо играет роль в образовании белков хлоропластов. При недостатке железа нет условий для образования таких важнейших компонентов хлоропластов, как цитохромы, ферредоксин и некоторые другие. Возможно, это косвенно влияет на образование хлорофилла. В хлоропластах железо в негемовой форме входит в состав реакционных центров фотосистем I и II.

