2013-12-31
2013-12-31 792
792
Стратегия фитоалексинной защиты растения может быть разной. В устойчивых к антракнозу (возбудитель Colletotrichwn lindemutianum) сортах фасоли ФА фазеоллин накапливается в высокой концентрации уже через 2...3 ч после заражения, в то время как в восприимчивых сортах такая же концентрация фазеоллина достигается значительно позднее— через 7...8ч и не препятствует развитию инфекции и образованию потомства возбудителя. Причина таких различий в чувствительности в разные периоды инфекционного процесса — неодинаковая чувствительность к ФА спор и мицелия гриба. Прорастание спор подавляется при концентрации 10 мкг/г, а удлинение ростковой трубки — еще при более низкой — 3 мкг/г. Поэтому быстрое накопление фазеоллина на ранних этапах инфекционного процесса останавливает развитие паразита. В то же время на рост гиф фазеоллин влияет значительно слабее — при концентрации 68 мкг/г рост лишь несколько тормозится, поэтому позднее накопление ФА было неэффективным. Иная стратегия защиты наблюдается у сои, зараженной Phytophthora sojae. Концентрация ФА глицеоллина начинает расти с одинаковой скоростью после заражения восприимчивых и устойчивых сортов, но у первых она, достигнув максимума через 5...6 ч, падает,'а у вторых продолжает расти, достигая значительно более высокого максимума через 24 ч после заражения.
защитные белки растений. Р R - б е л к и. Белки, связанные с патогенезом (PR-белки), образуются в растениях в ответ на заражение грибными," бактериальными, вирусными болезнями, а также в ответ на обработку некоторыми химикатами. Все эти белки обладают широким антигрибным спектром действия. Среди них большой интерес представляют следующие.
• Ферменты р-1,3-глюканазы и хитиназы (группы PR-2 и PR-3 соответственно), разлагающие полисахариды р-1,3-глю-кан и хитин. Поскольку глюканы содержатся в клеточных стенках растений, наличие у них глюканаз как регуляторов процессов синтеза и деградации элементов клеточной стенки неудивительно. Но хитина у растений нет, а это означает, что хитиназа несет главным образом защитные функции. Синтез этих ферментов в ответ на заражение также свидетельствует об их связях с защитными реакциями. Хитин и (3-глюкан — основные компоненты клеточных оболочек грибов, из хитина построены стилеты сосущих насекомых и нематод. В настоящее время гены, ответственные за синтез хитиназы и Р -глюканазы, широко используются в генно-инженерных конструкциях с целью создания трансгенных растений, устойчивых к болезням и вредителям.
• Белки, связывающие хитин (группа PR-4). Они относятся к большой группе связывающих хитин белков — лектинов.
• Тауматиноподобные белки (группа PR-5), которые имеют структурное сходство со сладким белком тауматином из южноафриканского кустарника Taumatococcus. Они вызывают лизис грибного мицелия вследствие связывания р-1,3-глюкана в клеточных стенках грибов (обладают Р -1,3-глюканазной активностью), инги-бируют а-амилазу насекомых и человеческий трипсин (обладают свойствами ингибиторов ферментов).
Ингибиторы протеиназ. Ингибиторы протеиназ (ИП) — это белки, образующие комплексы с протеиназами и ингибирую-щие их каталитическую активность. Известно несколько классов ИП, отличающихся ингибированием разных типов протеиназ. Некоторые ИП накапливаются в незараженных растениях, синтез других индуцируется в ответ на заражение или повреждение насекомыми. ИП подавляют активность протеиназ, выделяемых в растение фитопатогенными грибами и насекомыми, и нарушают способность патогенов поглощать растительные белки, то есть снижают их пищевую активность. ИП не только выполняют защитные функции, они могут служить запасающими белками (накапливаются в покоящихся органах растений — семенах, клубнях) и регуляторами протеиназной активности самих растений. '
Антивирусные белки. У многих растений (фитоляки, дурмана, табака и др.) обнаружены белки, ингибирующие вирусную инфекцию. Эти белки разделяют на две большие группы: эндогенные, присутствующие в клетках независимо от заражения, и индуцированные, возникающие при заражении вирусами. Первые ингибируют главным образом ранние фазы вирусной инфекции путем агрегации ингибиторного белка с вирионом и инги-бирования репликации вирусных белков вследствие инактивации рибосом. Индуцированные антивирусные белки ингибируют репликацию вирусов. Некоторые из них имеют много общего с ин-терферонами человека (интерфероны ингибируют репликацию некоторых вирусов растений). Большинство антивирусных белков синтезируется в растении в очень низких концентрациях, что затрудняет их исследование.
Дефензины. Это низкомолекулярные (45...54 аминокислоты) белки, богатые цистеином. Дефензины продуцируются как защитные белки млекопитающими, насекомыми, грибами и растениями. Растительные дефензины могут вызывать морфологические изменения у восприимчивых видов грибов (морфоген-ные дефензины), ингибировать рост мицелия без морфологических изменений (неморфогенные дефензины), ингибировать а-амилазу грибов. Они связываются с рецепторами на мембранах грибов и вызывают усиление потери К+ и поступления Са2+ в клетки.
Другие защитные белки растений. Помимо дефензи-нов факторами устойчивости к патогенам могут быть и другие низкомолекулярные антимикробные белки, богатые цистеином. Это белки, осуществляющие трансмембранный транспорт л и п и д о в, белки, подавляющие синтез белка на чужеродных рибосомах на стадии элонгации (RIP-белки), и богатые серой ти -онины, накапливающиеся в семенах и проростках растений. Последние взаимодействуют с мембранными фосфолипидами и вызывают образование пор в мембранах. Эти белки обнаружены в семенах, листьях, цветках и клубнях разных растений, часто накапливаются в клеточных стенках, осуществляя защитную функцию.
пищевая недостаточность растений. Некоторые фитопато-генные грибы не способны синтезировать отдельные органические соединения (ауксотрофны по ним) и для своего роста должны получать эти соединения из растений-хозяев. Растения или их органы, в которых эти соединения отсутствуют или содержатся в низкой концентрации, устойчивы к данному паразиту или слабо поражаются им. Например, рост на питательной среде возбудителя фузариоза колоса {Fusarium graminearum) резко стимулируется азотсодержащими соединениями холином и бетаином. У пшеницы больше всего холина в пыльниках и других элементах цветков и очень мало в листьях и корнях, что и обусловливает органотропную специализацию гриба. Для роста паразита сливы Rhodostricta quercina требуется липоинозит, содержание которого у разных видов сливы коррелирует с их пораженностью.
Отмечены случаи активной реакции растения — снижение в нем синтеза необходимых паразиту продуктов в ответ на заражение. В лаборатории известного патофизиолога К. Т. Сухорукова было показано, что возбудитель вилта хлопчатника (Verticillium dahliae) нуждается в витамине биотине, причем его содержание в восприимчивых сортах хлопчатника после заражения возрастает, а в устойчивых — падает. Устранение необходимых паразиту веществ из очага поражения – одна из форм защитных реакций растений.
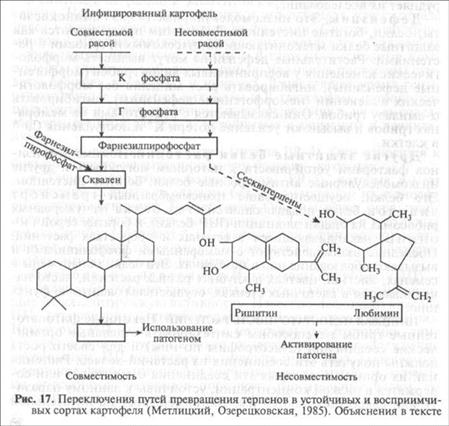
Паразиты из семейства Pythiaceae (роды Pythium и Phytophthora) не синтезируют фермент скваленсинтетазу и поэтому не образуют стеринов, которые необходимы им для формирования бесполого и полового спороношения, а также для патогенное™. Но они используют фитостерины растений-хозяев, трансформируя их в нужные для себя вещества. Заражение картофеля возбудителем фитофтороза активизирует синтез тер-пеноидного метаболизма и накопление в зоне заражения нужных паразиту стеринов. Как установили биохимики Л. В. Метлицкий и О. Л. Озерецковская, синтез терпеноидов в устойчивых и восприимчивых сортах картофеля идет одинаково до стадии образования С15 фарнезилпирофосфата, однако далее в восприимчивых сортах под действием фермента скваленсинте-тазы две молекулы фарнезилпирофосфата конденсируются с образованием С3о сквалена, из которого образуются фитостерины, а в устойчивых — под действием другого фермента (циклазы) молекулы фарнезилпирофосфата замыкаются в два кольца и образуются бициклические сесквитерпены, являющиеся высокотоксичными фитоалексинами (см. рис. 16). Таким образом, вместо необходимого питательного вещества растение подсовывает паразиту токсин (рис. 17).