2013-12-31
2013-12-31 900
9001. Ахарактарызуйце аддзел мохападобных.
2. Назавіце рысы прымітыўнай арганізацыі пячоначных імхоў.
3. Дайце характарыстыку сфагнавых імхоў.
4. Якія агульныя прыкметах сфагнавых і андрэевых імхоў?
5. Якая будова спарагона зязюліна льну звычайнага?
6. Як тлумачыцца паходжанне мхоў?
Дыплоідная лінія эвалюцыі вышэйшых раслін
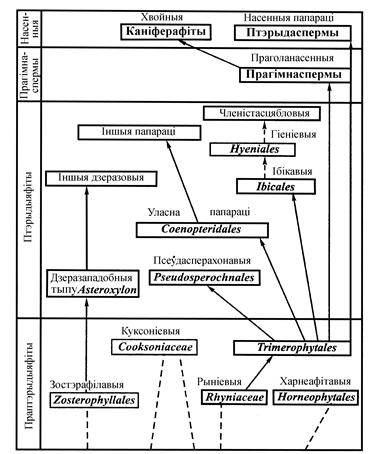
АДДЗЕЛ РЫНІЯФІТЫ (RHYNIOPHYTA)
Гэты аддзел аб‘ядноўвае вымерлыя старажытнейшыя вышэйшыя расліны, вядомыя з сілуру да верхняга дэвону (435-345 млн. гадоў назад, гл. табліцу 2).
Упершыню рэшткі рыніяфітаў знойдзены канадскім геолагам Дж. Досанам у дэвонскіх адкладаннях (гл. табліцу 2) паўвострава Гаспэ (Канада) у 1859 г. Досан назваў гэту расліну Psilophyton princeps.
Пазней, у 1917 г., англійскія палеабатанікі Р. Кідстан і У. Ланг апісалі рэшткі раслін з адкладанняў крэмніевай пароды Шатландыі дэвонскага
ўзросту – псілафіт, рынію, харнеафітон і іншых (малюнак 18). Яны вызначылі роднасць іх з раней адкрытым Psilophyton princeps і аб’ядналі іх у новы парадак псілафітавыя (Psilophytales).
У 1937 г. У. Ланг адкрыў новы род – куксонію (Сooksonia). Гэта расліна была знойдзена таксама ў верхнесілурыйскіх адкладаннях Чэхіі, Славакіі, краін СНД, ЗША. Яна жыла каля каля 415 млн. гадоў таму назад (гл. табліцу 2).
Табліца 2 – Паслядоўнасць геалагічных перыядаў і развіцця расліннага свету
| Эры | Перыяды | Радыялагічнае датаванне, млн. гадоў | Пануючыя групы раслін | |
| Працягласць | Узрост ніжняй мяжы | |||
| Кайназой | Чацвярцічны | Панаванне сучасных пакрытанасенных з перавагай траў | ||
| Трэцічны | 67 | Панаванне пакрытанасенных з перавагай дрэў | ||
| Мезазой | Мелавы | З’яўленне пакрытанасенных, выміранне сагаўнікаў і гінкгавых | ||
| Юрскі | Панаванне голанасенных (сагаўнікаў, гінкгавых, хвойных) | |||
| Трыясавы | Развіццё голанасенных. Выміранне кардаітаў |
Заканчэне табліцы 2
| Палеазой | Пермскі | Выміранне дрэвападобных дзеразовых, хвашчовых. З’яўленне насенных папарацей | ||
| Каменна-вугальны | Развіццё папарацепадобных (папараці, хвашчы, дрэвападобныя дзярэзы), кардаітаў | |||
| Дэвонскі | Развіццё рыніяфітаў, дзеразовых, хвашчоў, папарацей. З’яўленне першых голанасенных | |||
| Сілурыйскі | З’яўленне першых наземных раслін – рыніяфітаў, імхоў | |||
| Ардовіцкі | Перавага марскіх водарасцей і бактэрый | |||
| Кембрыйскі | ||||
| Пратэ-разой | Вендскі | Нямногія прыкметы водарасцей,бактерый | ||
| Рыфейскі | ||||
| Афебійскі | ||||
| Архей | 1500-5000 | Вапнякі, б.-м. бактэрыяльнага паходжання |
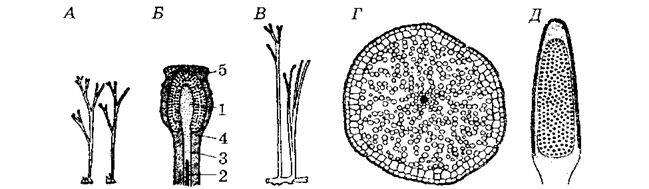
Рынія, харнеяфітон (малюнак 18), куксонія, псілафіт і інш. складаюць найбольш старажытны і самы прымітыўны аддзел вышэйшых раслін, геалагічная гісторыя якіх пачынаецца з сілурыйскага перыяду і да верхняга дэвону (гл. табліцу 2). Яго называюць рыніяфіты (Rhyniophyta).
Гэта найбольш проста арганізаваная група вышэйшых выкапнёвых раслін (малюнак 18). У іх быў карэнішчападобны орган рызамоід (ад грэч. rhiza – корань), ад якога ўверх адыходзілі тэломы (ад грэч. telos – канец) – першасныя надземныя цыліндрычныя органы вышэйшых раслін. На вяршынях некаторых тэломаў размяшчаліся прадаўгаватыя таўстасценныя спарангіі. Участкі паміж разгалінаваннямі тэломаў атрымалі назву месомаў (ад грэч. mesos – сярэдні). Лісты і карані ў іх адсутнічалі. Анатамічная будова рэніяфітаў вельмі прымітыўная. Праводзячая сістэма была слаба развіта і ўяўляла сабой пратастэлу. Наяўнасць покрыўных тканак і вусцейкаўсведчыць пра тое, што гэта былі наземныя ці каляводныя расліны. Іх памеры дасягалі 50см вышыні і каля 5мм у дыяметры, але расліны сярэднедэвонскага роду Pertiса мелі вышыню ў 3м (малюнак 20).
Мяркуемыя гаметафіты харнеафітона апісаны пад назвай ліёнафітон (Lyonophyton). Іх восі мелі анатамічную будову, падобную з харнеяфітонам. На вяршыні некаторых тэломаў ліёнафітона былі чашабадобныя пашырэнні з лопасцевымі краямі. У цэнтры чаша ўтварала выступ, у які былі пагружаны архегоніі з моцна адцягнутай шыйкай. Па перыфірыі чашы, з яе ўнутранага боку, размяшчаліся акруглыя антэрыдыі.
У іншых прадстаўнікоў рыніяфітавых гаметафіты не апісаны.

Класіфікацыя раніяфітаў не распрацавана. Вылучаюць клас Rhyniopsіda, парадкі Rhyniales і Psilophytales, ці Trimerophytales.
Рыніяфіты далі пачатак макрафільнай лініі эвалюцыі вышэйшых раслін. З імі філагенітычна звязаны псілотападобныя, хвошчападобныя, а праз трымерафітавых - з папарацепадобнымі і прагімнаспермамі.
Прыхільнікі рыніяфітнай гіпотэзы паходжання мохападобных (Р. Кідстан, У. Ланг, Т. Хале, Г. Эндрус, Б. М. Коза-Палянскі) накарысць гэтай гіпотэзы прыводзяць некаторыя палеабатанічныя дадзеныя. У харнеяфітона (Horneophyton) з рыніяфітаў у спарангію ёсць калонка, падобная на калонку ў каробачцы сфагнуму і андрэа. Дэвонскі род Sporogonites мае спарафіт у выглядзе доўгай ножкі са спарангіем, у яго ёсць калонка, як у сфагнавых імхоў.
Адкрыццё і вывучэнне рыніяфітаў, палажэнняў, выказаных у працах Патанье, Лінга і Баўэра паслужылі асновай для стварэння тэломнай тэорыі, аўтарам якой быў нямецкі вучоны В.Цымерман (1930 г.). Гэтая тэорыя раскрывала асаблівасці будовы першых наземных раслін і паказвала магчымыя шляхі, па якіх адбывалася станаўленне асноўных вегетатыўных і рэпрадуктыўных органаў вышэйшых раслін розных сістэматычных груп.
Адпаведна ўяўленням Цымермана, цела першых наземных раслін складалася з радыяльных сіметрычных восей. Іх канечныя галінкі атрымалі назву тэломаў. Тэломы галінаваліся дыхатамічна ва ўзаемна перпендыкулярных плоскацях, утваралі аб’ёмную сістэму тэломаў (малюнак 19).
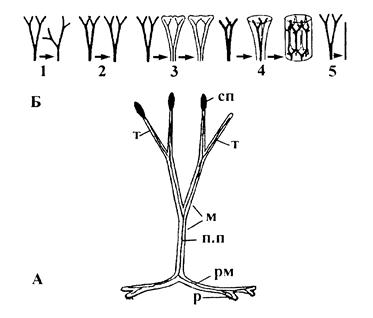

Па меры разгалінавання тэломы з канечных становяцца прамежкавымі, з-за чаго атрымалі назву месомаў. Адметнай асаблівасцю тэломаў была наяўнасць праводзячых пучкоў, пабудаваных па тыпу пратастэлы. Гэтым яны прынцыпова адрозніваліся ад водарасцей. Падземныя, ці сцелістыя, тэломы атрымалі назву рызамоідаў, ці карэнішчападобных. У ходзе развіцця рыніяфітаў тэломы падвергнуліся шэрагу змяненняў і ў выніку ператварыліся ў асноўныя органы вышэйшых раслін – сцёблы, лісты, карані, спарафілы. Пры гэтым адбыліся эвалюцыйныя працэсы (малюнак 19Б): перавяршыньване, планацыя, зрастанне ў адной плоскасці, зрастанне ў розных плоскасцях, рэдукцыя.
Перавяршыньванне адбывалася ў выніку няроўнай дыхатаміі, што добра прасочваецца ў рыніяфітаў. У выніку сфарміраваліся галоўная вось і сістэма бакавых галін. Гэта з’явілася перадумовай для стварэня буйных формаў раслін.
Пад планацыяй разумеецца размяшчэнне тэломаў у адной плоскасці. Гэты працэс з’явіўся важнейшым этапам у ходзе фарміравання лістоў папарацей, хвошчападобных, праголанасенных раслін. Зрастанне тэломаў як канчатковы вынік філагенетычнага працэсу мог распаўсюджвацца на тэломы, размешчаныя як у адной, так і ў некалькіх плоскасцях. Калі зрастанне спалучалася з планацыяй, то фарміраваліся буйныя рассечаныя лісты з сістэмай разгалінаваных жылак. Зрастанне тэломаў, размешчаных у розных плоскасцях, прыводзіла да ўтварэння сцёблаў са стэламі складанай канфігурацыі (напрыклад, актынастэла ў астэраксілона).
Пад рэдукцыяй разумеецца памяншэнне ліку парадкаў разгалінавання, што прыводзіць да спрашчэння будовы органа. Так, простая ліставая пласцінка хвашчоў у ходзе эвалюцыі ўзнікла з сістэмы шматразова разгаліноўваемых тэломаў, якія падвергліся рэдукцыі. Прынцыпова важным палажэннем тэломнай тэорыі з’яўляецца тое, што ўзнікненне вегетатыўных лістоў і спарафілаў адбывалася адначасова і незалежна адно ад другога, і далейшае іх развіццё ішло паралельнымі шляхамі. Такім чынам, фарміраванне вегетатыўных і спараносных парасткаў адбывалася таксама незалежна адно ад другога.
Аналіз палеабатанічнага матэрыялу і ход развіцця сучасных прымітыўных раслін паказвае, што калі не ва ўсіх, то ў большасці вышэйшых раслін ход станаўлення вегетатыўных і спараносных органаў адпавядае асноўным палажэнням таломнай тэорыі.
АДДЗЕЛ ЗОСТЭРАФІЛАФІТЫ (ZOSTEROPHYLLOPHYTA)
Зостэрафілафіты аб’ядноўваюць невялікую групу ранне- і сярэднедэвонскіх раслін. Яны маюць шмат агульнага з рыніяфітамі. Іх нават уключалі ў аддзел рыніяфітаў, у ранг самастойнага класа зостэрафілапсіды (Zosterophyllopsida). Некаторыя батанікі збліжаюць зостэрафілапсіды з дзеразападобнымі і нават аб’ядноўваюць іх па прыкмеце не верхавіннага, а бакавога размяшчэння спарангіяў і экзархнай ксілемай (найбольш старыя элементы ксілемы максімальна аддалены ад цэнтра органа). Але ад дзеразападобных зостэрафілафіты рэзка адрозніваюцца адсутнасцю лісцевых органаў – асаблівасцю, якая характэрна толькі для рыніяфітаў. Таму Д. Бірхарст (1971) вылучае гэтую групу вымерлых раслін у самастойны аддзел. Ён выказвае меркаванне, што зостэрафілафіты былі той групай, якая паклала пачатак аддзелу дзеразападобных. Аднак не выключаецца і тое, што зостэрафілафіты ўяўляюць сабой толькі бакавую галіну эвалюцыі, якая ўзнікла ад агульнага з дзеразападобнымі продка. Не выключана, што зостэрафілафіты і дзеразападобныя ўзніклі ад агульнага продка, які ўзяў пачатак ад рыніяфітаў.
У аддзел зостэрафілафітаў уваходзіць клас зостэрафілапсіды (Zosterophyllopsida), парадак зостэрафілавыя (Zosterophyllales) і дзве сям’і: Zosterophyllaceae і Gosslingiaceae.
Выкапнёвыя трымерафітавыя і зостэрафілавыя паказаны на малюнку 20.
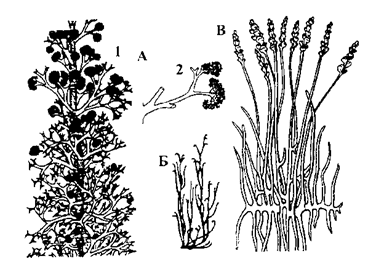

Род зостэрафілум (Zosterophyllum) адкрыты ў 1892 годзе. Вядомы з ніжняга дэвону. Ён быў шырока распаўсюджаны ў Еўропе, Заходняй Сібіры, Кітаі, Паўночнай Амерыцы, Аўстраліі. Гэта былі маленькія, вышынёй 15-17см, дыхатамічна разгалінаваныя наземныя расліны. Спарангіі размяшчаліся на кароткіх бакавых галінках ці былі сабраны ў коласападобнае ўтварэнне ў верхняй частцы тэломаў.
Цікавы род саўдонія (Savdonia), восі якой мелі шыпаватыя вырасты эмергенцыі. На восях размяшчаліся спарангіі на вельмі кароткіх і тоўстых ножках (малюнак 20 Б).
Роды гаслінгія (Gosslingia) і крэнатакаўліс (Crenatocaulis) аб’ядноўваюць у сям’ю гаслінгіевыя (Gosslingіaceae).
Філагенетычныя сувязі рыніевых і зостэрафілавых (па С. В. Мейену) прыведзены на схеме (малюнак 21).
АДДЗЕЛ ДЗЕРАЗАПАДОБНЫЯ (LYCOPODIOPHYTA)
Дзеразападобныя, як і мхі, ўяўляюць сабой адну з наібольш старажытных груп вышэйшых раслін. Яны вядомых з сілуру як дробныя травяністыя, так і дрэвападобныя формы. Амаль дакладна можна лічыць продкамі дзеразападобных зостэрафілафіты. Найбольшага развіцця дзеразападобныя дасягнулі ў карбоне (гл. табліцу 2). У цяперашні час гэты аддзел прадстаўлены невялікай колькасцю родаў і відаў. Сучасныя прадстаўнікі – шматгадовыя травяністыя расліны, звычайна вечназялёныя. Сярод выкапнёвых былі як травяністыя, так і магутныя дрэвавыя формы. Дзеразападобныя прадстаўлены як раўнаспоравымі, так і разнаспоравымі мікрафільнымі раслінамі. Гэта характэрныя прыкметы аддзела.
Гаметафіты раўнаспоравых сучасных дзеразападобных маюць памер ад 2 да 20мм. Вядуць мікатрофны лад жыцця. Паспяваюць на працягу 1-15 гадоў. Гаметафіты разнаспоравых дзеразападобных аднаполыя, развіваюцца на працягу некалькіх тыдняў за кошт спажыўных рэчываў, якія ўтрымліваюцца ў споры.
Жаночы гаметафіт утвараецца пры прарастанні мегаспоры, разрывае яе абалонку, выпукваецца вонкі, клеткі яго зелянеюць, утвараюцца рызоіды. Ён вядзе самастойны лад жыцця. На ім развіваецца некалькі архегоніяў з яйцаклеткамі. Мікраспора пры прарастанні ўтварае моцна рэдукаваны мужчынскі гаметафіт з антэрыдыем. У антэрыдыю фарміруюцца двухжгуцікавыя сперматазоіды. Апладненне адбываецца ў кропельна-вадкім асяроддзі.
Аддзел уключае два класы: дзеразовыя (Lycopodiopsida),раўнаспоравыя іселагінелавыя (Selaginellopsida), у асноўным разнаспоравыя.
КЛАС ДЗЕРАЗОВЫЯ (LYCOPODIOPSIDA)
У клас дзеразовыя ўваходзяць як вымерлыя прадстаўнікі парадкаў астэраксілавых (Asteroxylales), дрэпанафікусавых (Drepanophycales), так і цяпер існуючыя – парадак дзеразовыя (Lycopodiales).
Парадак астэраксілавыя (Аsteroxylales)
Парадак утрымлівае адну сям’юастэраксілавыя (Asteroxylaceae) і два роды– астэраксілон (Asteroxylon) і схізаподыум (Schisopodium). Абодва роды вымерлі к канцу дэвону (360-370 млн. гадоў назад). Вядомы з сярэдняга дэвону (435 млн. гадоў назад).
Астэраксілон (малюнак22А) быў травяністай раслінай, з надземнай і падземнай часткамі, сцяблом і лістамі. Лісты ўяўлялі сабой вырасты сцябла (энацыі). Верагодна, астэраксілон даў пачатак вышэйшым раслінам з дробнымі лістамі – мікрафільнай лініі ў эвалюцыі вышэйшых раслін. Праводзячая сістэма астэраксілона – актынастэла.
Парадак дрэпанафікусавыя (Drepanophycales)
Парадак аб’ядноўвае 2 роды і 7 відаў.
Род дрэпанафікус (Drepanophycus, малюнак 22Б)вядомы з ніжняга,сярэдняга і верхняга дэвону ўсіх мацерыкоў зямнога шара. Гэта былі расліны да 50см вышынёй, мелі сцелістыя і вертыкальныя, амаль дыхатамічна разгалінаваныя парасткі. Вузкія лісцікі мелі адну жылку. Спарангіі размяшчаліся або ў похвах лістоў, або на паверхні спарафілаў. Стробілаў яшчэ не было.
Род барагванацыя (Baragwanathia, малюнак 22В) – з ніжняга дэвону. Дыхатамічна разгалінаваныя парасткі былі пакрыты вузкімі (да 1 мм) і адносна доўгімі (да 4 см) лістамі. Спарангіі ныркападобнай формы размяшчаліся на верхняй паверхні спарафілаў. Верагодна, гэтыя дэвонскія расліны ўяўлялі сабой бакавую галіну эвалюцыі дзеразападобных.


У раслін парадку дрэпанафікусавыя праявіліся тры асноўныя тэндэнцыі развіцця дзеразападобных: 1) васкулярызацыя лістоў; 2) перамяшчэнне спарангіяў у пазуху ліста, а ў далейшым і на яго паверхню; 3) пераход ад беспарадкавага размяшчэння спарафілаў да фарміравання спараноснай зоны, а ў далейшым – да спараноснага парастка – стробіла. Дрэпанафікусавыя і блізкія да іх вымерлыя роды далі пачатак тром асноўным філагенетычным лініям дзеразападобных – травяністым раўнаспоравым (парадак дзеразовыя), травяністым разнаспоравым (парадак селагінелавыя) і вымерлым дрэвавым разнаспоравым (парадак лепідадэндравыя).
Парадак дзеразовыя (Lycopodiales)
У яго ўваходзіць дзве сям’і дзеразовыя (Lycopodiaceae) і баранцовыя (Huperciactae) і ад 2 да 6 родаў. Род філаглосум (Phylloglossum) сустракаецца ў Аўстраліі.
Род дзераза налічвае ад 200 да 500 відаў. У краінах СНД прадстаўлены 10-12 відамі, у Рэспубліцы Беларусь – 6 відамі.
Сучасная дзераза – невялікая травяністая расліна. У трапічных лясах сустракаецца Lycopodium volubile, дасягае 9м удаўжыню (ліяна).
У нашай краіне шырока распаўсюджана дзераза булавападобная (Lуcopodium clavatum, малюнак 23), якая расце ў хвойных, часцей сасновых лясах. Дыхатамічна разгалінаванае сцябло дзеразы сцелецца па зямлі, чапляецца, дзярэцца. Адсюль, мабыць, і назва дзеразы.
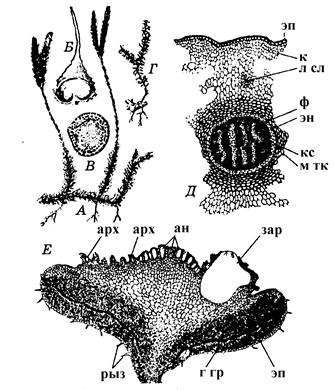

Анатамічная будова сцябла прымітыўная. Маладыя сцяблінкі маюць пратастэлічную, а дарослыя – плектастэлічную будову. Звонку сцябло пакрыта эпідэрмай. Далей ідзе моцная кара. Клеткі ўнутраных слаёў кары маюць тоўстыя сценкі і ўтвараюць механічнае кальцо. Цэнтральная частка сцябла занята канцэнтрычным праводзячым пучком (стэлай). Ксілема ў пучку размяшчаецца ў выглядзе стужак, якія злучаны паміж сабой. Паміж участкамі ксілемы і вакол яе з перыферыі - флаэма. Асяродка няма, камбія ў пучку няма. За флаэмай ідзе слой перыцыкла, далей – эндадэрма, якая аддзяляе пучок ад кары. У кары сцябла ёсць ліставыя сляды – пучкі праводзячай тканкі, якія ідуць да жылак ліста.
Лісты звонку адзеты эпідэрмісам з вусцейкамі. Мезафіл шматслойны з міжклетнікамі. Карані бакавыя з каранёвымі валаскамі, жывуць 2-5 гадоў.
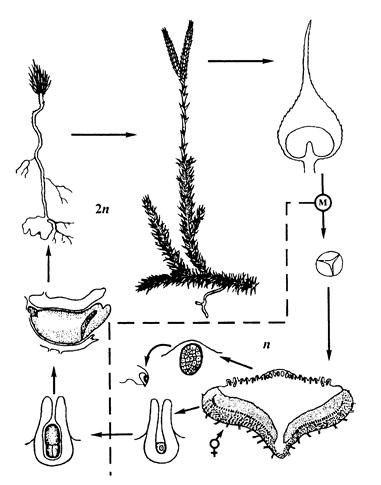
Спаранашэнне ў дзеразы назіраецца ў канцы лета-восенню. Спараносныя каласкі па адным, радзей па два-тры ўзнікаюць на верхавінах галінак. На восі каласка размешчаны спарафілы. Яны драбнейшыя за вегетатыўныя лісты, маюць трохвугольна-яйцападобную форму, завостраныя, жоўтага колеру.
На верхнім баку спарафілаў развіваюцца спарангіі ныркападобнай формы на кароткай ножцы. Унутры спарангія са спарагеннай тканкі шляхам меёзу фарміруюцца дробныя аднолькавыя споры ў вялікай колькасці. Яны маюць акругла-тэтраэдрычную форму жаўтаватага колеру. Споры маюць дзве абалонкі – экзаспорый і эндаспорый. Унутранае змесціва споры складаецца з пратаплазмы, ядра, пластыд, кропелек тлушчу. Спелы спарангій лопаецца папярочнай трэшчынай. Да гэтага часу вось каласка некалькі разрастаецца, спарафілы рассоўваюцца і споры лёгка высяваюцца на зямлю.
На яе паверхні споры дажджавой вадой заносяцца ў паглыбленні (шчыліны) глебы, дзе на глыбіні ў некалькі сантыметраў прарастаюць. Утвараецца зарастак у выглядзе камячка клетак. Жыццё зарастка магчыма толькі пры сімбіёзе з гіфамі грыба.
Спелы зарастак мае форму дзіцячай цацкі ваўчка. На верхнім баку зарастка ўтвараюцца антэрыдыі і архегоніі. Антэрыдыі пагружаны ў тканку зарастка. У іх развіваюцца шматлікія двухжгуцікавыя сперматазоіды. Архегоніі таксама пагружаны ў зарастак брушкам, а шыйка ўзвышаецца над паверхняй зарастка. У брушку архегонія фарміруецца яйцаклетка і брушная канальцавая клетка. У шыйцы – 6-8 канальцавых клетак. Апладненне адбываецца ў кропельна-вадкім асяроддзі.
Аплодненая яйцаклетка дзеліцца і фарміруе зародак, які мае сцяблінку, першасны лісцік і бакавы корань. На першых этапах зародак жывіцца за кошт спажыўных рэчываў зарастка. Зарасткі дзеразы развіваюцца вельмі марудна. Ад прарастання споры да ўтварэння спарафіта праходзіць да 15-16 гадоў.
Цыкл развіцця дзеразы булавападобнай паказаны на малюнку 23.
Рысы прымітыўнай арганізацыі дзеразы: 1) дыхатамічнае разгалінаванне сцяблоў і каранёў; 2) адсутнасць строгай дэтэрмінаванасці пры закладванні парасткаў і каранёў; 3) бакавыя карані; 4) анатамічнае падабенства ў будове сцяблоў і каранёў; 5) наяўнасць самых прымітыўных тыпаў стэлы ў шэрагу відаў – пратастэлы, плектастэлы і актынастэлы; 6) вялікая працягласць жыцця гаметафітаў; 7) наяўнасць доўгай шыйкі архегонія. Усё гэта дае падставу разглядаць віды родаў дзеразовых як сапраўды жывых выкапнёвых раслін.

Дзеразовыя маюць гаспадарчае выкарыстанне. Споры дзеразы ў медыцыне вядомы як дзіцячая прысыпка. Імі таксама перасыпаюць пілюлі. У металургіі пры адліўцы больш дакладных дэталей зямельныя формы пасыпаюць спорамі дзеразы. У сельскай мясцовасці нярэдка дзеразой абвіваюць аркі, партрэты, што рабіць немэтазгодна па прычыне зберажэння гэтай расліны.
У Чырвоную кнігу Рэспублікі Беларусь з дзеразападобных занесены два віды: дзераза баранец звычайны (Huperzia selago) і лікападыела заліваемая (Lycopodiella inundata).
КЛАС СЕЛАГІНЕЛАВЫЯ (SELAGINELLOPSIDA)
Да гэтага класа адносяцца як выкапнёвыя, так і цяпер жывучыя разнаспоравыя шматгадовыя травяністыя расліны (парадкі селагінелавыя – Selaginellales і палушнікавыя – Isoёtales), а таксама пераходныя ад травяністых да драўняных разнаспоравых раслін (парадкі проталепідадэндравыя – Protolepidodendrales і лепідадэндравыя – Lepidodendrales). У пазусе ліста селагінелавых маецца язычок – лігула.
Парадак селагінелавыя (Selaginellales)
Да гэтага парадку адносяцца разнаспоравыя травяністыя расліны, якія бяруць пачатак ад дрэпанафікусавых ці проталепідадэндравых. Парадак уключае адну сям’ю селагінелавыя (Selaginellaceae) з адным родам селагінела(Selaginella, малюнак 25). Ён утрымлівае да 700 відаў, якія растуць у вільготных лясах тропікаў і субтропікаў. Нямногія віды сустракаюцца ва ўмераных шыротах. У краінах СНД сустракаецца 8 відаў селагінелы, у Беларусі яны адсутнічаюць.
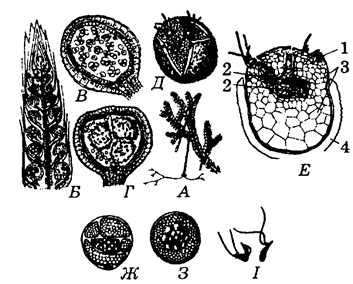

У выкапнёвым стане селагінелавыя сустракаюцца з позняга дэвону і ранняга карбону.
Селагінелы даволі разнастайныя па знешнім выглядзе. Сярод іх ёсць як паўзучыя, так і вертыкальныя формы. Памер іх ад 5-10см да 2-20м удаўжыню. У тропіках некаторыя з іх пакрываюць глебу лясоў, як імхі і лішайнікі ў нашых лясах.
Селагінела селагападобная (S. selaginoides) сустракаецца ў гарах Каўказа, Урала, Сібіры, Далёкага Усходу. Гэта шматгадовая травяністая расліна памерам каля 10см. У яе, як і ў дзеразы булавападобнай, сцябло дыхатамічна разгалінавана, лісты размешчаны ў два рады. Ніжні рад утвораны лістамі больш шырокімі, гарызантальна арыентаванымі да паверхні глебы. Верхні рад лістоў прадстаўлены меншымі лісточкамі, якія не засланяюць ніжні рад ад сонечнага святла. Ад сцябла адыходзяць караняносцы (рызафоры), ад іх – дыхатамічна разгалінаваныя карані.
Сцябло селагінелы звонку пакрыта аднаслойнай эпідэрмай без вусцеек. Далей ідзе моцная кара, знешні слой якой з таўстасценных клетак, выконвае механічную функцыю. У цэнтры, у паветранай поласці падвешаны пры дапамозе аднародных зялёных клетак трабекулярных ніцей (клеткі эндадэрмы) праводзячы пучок (стэла). Лісты маюць жылку пасярэдзіне. У пазусе ліста маецца язычок (лігула). Гэта важная сістэматычная прыкмета класа. Звонку ліст пакрыты эпідэрмай. У клетках ліста – адзін-два пласціністых храматафоры, як у водарасцей. Вусцейкі размешчаны на ніжнім баку ліста. У аснове ліста знаходзіцца група рыхлых клетак (падушачка), да якой падыходзіць трабекулярная тканка, праз якую адбываецца паступленне паветра ў паветравую поласць сцябла.
Селагінела – разнаспоравая расліна. У яе фарміруюцца мікраспарангіі і мегаспарангіі. У мікраспарангіях шляхам рэдукцыйнага дзялення развіваецца вялікая колькасць мікраспор. У мегаспарангіях, таксама ў выніку рэдукцыйнага дзялення, утвараецца па 4 буйныя мегаспоры.
Спарангіі размяшчаюцца на верхнім баку спарафілаў, якія ўтвараюць спараносны каласок.
Мікра- і мегаспарангіі сабраны большай часткай у адным каласку. У такім выпадку мікраспарангіі знаходзяцца ў верхняй частцы ці ў другім радзе аднаго і таго ж каласка, а мегаспарангіі размешчаны ў ніжняй частцы каласка ці займаюць другі рад.
Пры прарастанні мікраспоры на зямлі ці ўнутры мікраспарангія ўтвараецца моцна рэдукаваны мужчынскі зарастак. Ён не пакідае абалонкі споры. Пры першым дзяленні мікраспоры ўтвараецца адна маленькая праталіяльная (рызаідальная) клетка і большая – антэрыдыяльная. У далейшым антэрыдыяльная клетка дае пачатак адзінаму антэрыдыю. Сперматагенныя клеткі ў выніку наступнага дзялення ўтвараюць вялікую колькасць двухжгуцікавых сперматазоідаў.
Мегаспора пры прарастанні ўтварае жаночы зарастак. Часцей гэта адбываецца на зямлі. Ядро мегаспоры шматразова дзеліцца, узнікаюць перагародкі і фарміруецца мнагаклетачны зарастак, які не пакідае мегаспоры. Клеткі яго ўзмоцнена дзеляцца і прарываюць абалонку мегаспоры. Зарастак выпучваецца вонкі, клеткі яго зелянеюць. На ім узнікаюць рызоіды, і ён вядзе самастойны лад жыцця. У верхняй частцы жаночага зарастка развіваецца некалькі архегоніяў, пагружаных брушкам у тканку зарастка. Апладненне адбываецца ў вадным асяроддзі.
Аплодненая яйцаклетка дзеліцца. Верхняя клетка ўтварае падвесак. З ніжняй фарміруецца зародак, які мае падвесак, верхавінную клетку, сцяблінку, зародкавыя лісточкі, ножку і рызафор. Пры дапамозе ножкі зародак усмоктвае спажыўныя рэчывы з зарастка. Пазней зародак пераходзіць да самастойнага жыцця і дае дарослую расліну селагінелы.
У некаторых відаў селагінелы (S. apus, S. rupestris) развіццё зарастка і апладненне адбываецца ўнутры мегаспарангія, на мацярынскай расліне. І толькі пасля гэтага мегаспарангій ападае. Тут да утварэння насення застаецца адзін крок.
Цыкл развіцця селагінелы прадстаўлены на малюнку 26.
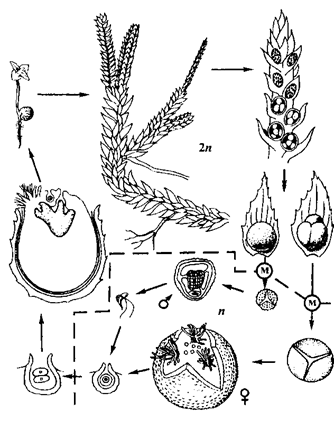

Разнаспоравасць у эвалюцыйным развіцці вышэйшых раслін мае вялікае значэнне, таму што вядзе за сабой утварэнне двух родаў зарасткаў (гаметафітаў) – мужчынскіх і жаночых. Пры гэтым рэдукцыя гаметафітаў дасягае значна большай ступені. Гэта адлюстроўвае агульную тэндэнцыю эвалюцыі вышэйшых раслін – узрастаючую рэдукцыю гаметафітаў і ўсё большае пераважанне спарафіта ў цыкле іх развіцця.
Парадак палушнікавыя (Isoёtales)
Да гэтага парадку адносяццца разнаспоравыя шматгадовыя травяністыя расліны, лісты якіх маюць добра выражаны язычок-лігулу. Парадак уключае сям‘ю палушнікавыя, 3 роды – натгарсціяна (Nathorstiana), стылітэс (Stylites) і палушнік (Isoёtes).
Натгарсціяна – вымерлы род, вядомы з ніжняга мелу. Гэта была травяністая расліна вышынёй 12-20см з мясістым прамастаячым сцяблом, на вяршыні якога размяшчалася шылападобнае лісце. У ніжняй частцы меліся тоўстыя кароткія сцігмарыі з каранямі, інакш нагадвала плеўрамею ў мініяцюры (гл. малюнак 27).
Бліжэй за ўсё да натгарсціянай стаіць род сцілітэс, адкрыты ў 1945 г. у цэнтральнаперуанскіх Андах на вышыні 3800 – 4750м над узроўнем мора. Гэтая расліны ад 7 да 20см увышыню, мела адносна тоўстае (да 3-х см у дыяметры), злёгку разгалінаванае сцябло напалову схаванае ў глебе. На верхавіне размяшчаўся пучок прадаўгавата-ланцэтных лістоў удаўжыню 5-6см. Ад асновы сцябла двума радамі адыходзілі мясістыя тоўстыя каранні. У сцябле выяўлены невялікі прырост другаснай ксілемы за кошт дзейнасці камбію (малюнак 28).
Найбольш шматлікі род палушнік (шыльнік ці ізает). Ён аб‘ядноўвае каля 70 відаў шматгадовых раслін, якія растуць пад вадой, альбо вядуць каляводны лад жыцця, толькі нямногія віды жывуць на моцна пераувільготненнай глебе.
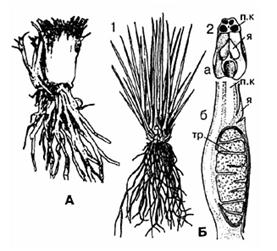

На ўнутранай пашыранай частцы спарафіла ў глыбокай ямцы знаходзяцца прыкрытыя язычком (лігулай) мега-і мікраспарангіі на кароткіх ножках ад 3мм да 3см. У мегаспарангіях развіваюцца ад 50 да 2000 мегаспор, а ў мікраспарангіях – ад некалькіх тысяч да соцен тысяч мікраспор.
Спарангіі паспяваюць, ападаюць, часта ўсплываюць на паверхню вады, згніваюць і выслабаняюць споры. Апошнія прарастаюць у гаметафіты.
Жаночы гаметафіт падобны на гаметафіт селагінелы, а мужчынскі мае будову, падобную на структуру гаметафіта селагінелы. Ён складаецца з адной праталіяльнай клеткі і аднаго антэрыдыя, які мае сценку з 4-х клетак і 2 сперматагенныя клеткі. Кожная сперматагенная клетка ўтварае па аднаму мнагажгутікаваму сперматазоіду.
Пасля апладнення з зіготы развіваецца зародак спарафіта, а затым – дарослая расліна.
Парадак проталепідадэндравыя (Protolepidоdendrales)
Парадак аб’ядноўвае 11 родаў і 50 відаў вымерлых палеазойскіх травяністых і драўняных раўнаспоравых раслін.
Самы старажытны родпроталепідадэндрон (Protolepidоdendron), вядомы з ніжняга і сярэдняга дэвону (гл. малюнак 22.Г). Ён быў увышыню 25 - 40см і адрозніваўся ад дрэпанафікуса пашыранай асновай ліста, дыхатамічным разгалінаваннем парасткаў і вяршынь некаторых лістоў. Спарангіі размяшчаліся на верхнім баку спарафілаў.
Расліны гэтага парадку ўяўляюць сабой пераходную групу ад травяністых дрэпанафікусавых, пазбаўленых ліставых падушачак, да дрэвавых разнаспоравых лепідадэндравых. Асаблівасць гэтага парадку – наяўнасць пашыраных асноў лістоў у выглядзе ліставых падушак.
Парадак лепідадэндравыя (Lepidоdendrales)
Гэты цалкам вымерлы парадак аб’ядноўвае 2 сям’і разнаспоравых дрэўных раслін. Вядомы і травяністыя формы (малюнак 28).
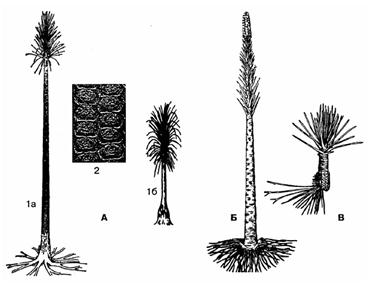
Малюнак 28 – Выкапнёвыя разнаспоравыя дзеразападобныя:
А – сiгiлярыя (Sigillaria): 1а – з карбона,
1б – з пермi, 2 – частка кары з падушачкамi; Б – плеўрамея (Pleuromeia); В – натгарсцiяна (Nathorstiana)
Палушнік, шыльнік (Isoёtes), (малюнак 29Б) распаўсюджаны на ўсіх кантынентах зямнога шара, у Беларусі расце палушнік азёрны (Isoёtes lacustris). Ён занесены ў нацыянальную Чырвоную кнігу. Гэтая травяністая шматгадовая расліна дасягае 8-25см увышыню. Сцябло палушніка кароткае, у аснове цыбулепадобна патоўшчанае, да асновы двух-трохлопасцевае. Ад сцябла паміж лопасцямі адыходзяць шматлікія карані. Вышэй на сцябле размешчаны лісты. Яны ўнізе расшыраныя, з добра выражаным язычком, а кверху звужаюцца і становяцца шылападобнымі (адсюль назва расліны – шыльнік). Унутры лістоў амаль па ўсёй даўжыні праходзяць паветраныя каналы. Пашыраная лопасцевая аснова сцябла адпавядае рызафору, а верхняя частка, ад якой адыходзяць лісты, – сцяблу. Унутры сцябла праходзіць цыліндрычная стэла. Яна пашыраецца данізу і ўтварае дзве ці тры лопасці, ад якіх адыходзяць пучкі, накіраваныя ў карані, а ад верхняй часткі стэлы – адгалінаванні ў лісты. Стэла акружана шырокім кальцом кары. У парэнхіме, што ахоплівае стэлу, маецца праслойка мерыстэмы, аналагічная камбію. Тут адрозніваюцца рызаморфныя і сцябловыя мерыстэмы. У выніку дзейнасці камбію сцябло палушніка нарастае ўтаўшчыню.
Знешнія лісты палушніка звычайна з‘яўляюцца мегаспарафіламі, а далей углыб размешчаны мікраспарафілы, і, нарэшце, унутраныя лісты – вегетатыўныя.
Малюнак 29 – Сучасныя палушнiкавыя:
А – сцiлiтэс (Stylites), знешнi выгляд; Б – палушнiк (Isoёtes): 1 – знешнi выгляд,
2а – папярочны разрэз праз аснову спарафiла, 2б – падоўжны разрэз праз аснову спарафiла (я – язычок, п.к – паветраныя камеры, тр – трабекулы)
На ўнутранай пашыранай частцы спарафіла ў глыбокай ямцы знаходзяцца прыкрытыя язычком (лігулай) мега-і мікраспарангіі на кароткіх ножках ад 3 мм да 3 см. У мегаспарангіях развіваюцца ад 50 да 2000 мегаспор, а ў мікраспарангіях – ад некалькіх тысяч да соцен тысяч мікраспор.
Спарангіі паспяваюць, ападаюць, часта ўсплываюць на паверхню вады, згніваюць і выслабаняюць споры. Апошнія прарастаюць у гаметафіты.
Жаночы гаметафіт падобны на гаметафіт селагінелы, а мужчынскі мае будову, падобную на структуру гаметафіта селагінелы. Ён складаецца з адной праталіяльнай клеткі і аднаго антэрыдыя, які мае сценку з 4-х клетак і 2 сперматагенныя клеткі. Кожная сперматагенная клетка ўтварае па аднаму мнагажгутікаваму сперматазоіду.
Пасля апладнення з зіготы развіваецца зародак спарафіта, а затым – дарослая расліна.
Парадак проталепідадэндравыя (Protolepidоdendrales)
Парадак аб’ядноўвае 11 родаў і 50 відаў вымерлых палеазойскіх травяністых і драўняных раўнаспоравых раслін.
Расліны гэтага парадку ўяўляюць сабой пераходную групу ад травяністых дрэпанафікусавых, пазбаўленых ліставых падушачак, да дрэвавых разнаспоравых лепідадэндравых. Асаблівасць гэтага парадку – наяўнасць пашыраных асноў лістоў у выглядзе ліставых падушак.
Самы старажытны род проталепідадэндрон (Protolepidоdendron), вядомы з сярэдняга дэвону (гл. малюнак 22Г). Ён быў увышыню не болей 25см – 40см і адрозніваўся ад дрэпанафікуса толькі пашыранай асновай ліста, дыхатамічным разгалінаваннем парасткаў і вяршынь некаторых лістоў. Спарангіі размяшчаліся на верхнім баку спарафілаў.
Парадак лепідадэндравыя (Lepidоdendrales)
Гэты цалкам вымерлы парадак аб’ядноўвае 2 сям’і разнаспоравых дрэўных раслін. Вядомы і травяністыя формы.
Сям’я лепідадэндравыя (Lepidоdendraceae) уключае верхнедэвонскія і карбонавыя расліны. У дэвонскім перыядзе яны былі нешматлікімі і не перавышалі 6-8 м увышыню, а ў каменнавугальным перыядзе па ліку відаў і па вялікіх памерах яны былі пераважальнымі і займалі вядучае становішча ў складзе расліннасці Зямлі. Асноўны род лепідадэндран (Lepidоdendron), ці лускадрэў, атрымаў назву па рэштках кары, пакрытай ліставымі падушкамі ромбападобнай і лускападобнай формы. Прадстаўнік – лепідадэндран.
Сям’я сігілярыевыя (Sigillariaceae). Большасць карбонавых прадстаўнікоў дасягалі ў вышыню да 20-30 м. Сваю назву атрымалі ад шасцівугольнай формы ліставых падушачак (ад грэч. sigilla – пячатка).
У верхнім палеазоі (350-280 млн. г. назад) дзеразападобныя дасягнулі свайго росквіту. З класа палушнікавых лепідадэндрон, ці лускадрэў (Lepidodendron), сігілярыя (Sigillaria), з позняга дэвону – ранняга карбону і плеўрамея (Pleuromeia) у мезазоі былі магутнымі дрэвамі (малюнак 26). Яны прынялі ўдзел ва ўтварэнні каменнага вугалю. У канцы палеазойскай эры і ў пачатку мезазойскай усе буйныя прадстаўнікі дзеразападобных вымерлі, уступілі месца больш канкурэнтназдольным голанасенным. Дробныя травяністыя дзеразападобныя здолелі перажыць не толькі эру панавання голанасенных, але і паспяхова прыстасавацца ў далейшым да жыцця ва ўмовах складана арганізаваных супольніцтваў з перавагай кветкавых раслін.
Падводзячы вынікі характарыстыкі аддзела дзеразападобных, неабходна падкрэсліць своеасаблівасць гэтай групы, якая праявілася: а) ва ўнікальнасці лістоў энацыйнага паходжання; б) у асаблівасці каранёў дзеразы, якія ўзнікаюць у апексе сцяблоў; в) у розных шляхах фарміравання жыццёвых форм. Ад першасна травяных дрэпанафікусавых эвалюцыя пайшла па двух
напрамках – да сучасных травяністых форм дзеразовых і селагінелавых і да дрэвавых унікальных форм парадку лепідадэндравых. Адны, вельмі спецыялізаваныя формы, вымерлі ў палеазоі, а другія прайшлі шлях рэдукцыі вегетатыўнага цела, сфарміравалі другасна травяныя формы парадкаў дзеразовыя і палушнікавыя.
У сваім паходжанні дзеразападобныя, безумоўна, звязаны з рыніяфітамі, ад якіх узніклі дрэпанафікусавыя раўнаспоравыя, магчыма, драўнінныя і разнаспоравыя травяністыя (парадак дрэпанафікусавыя) і травяністыя раўнаспоравыя расліны (парадак астэраксілавыя). Раўнаспоравыя травяністыя дрэпанафікусавыя далі пачатак тром асноўным філагенетычным лініям дзеразападобных – раўнаспоравым першасна травяністым (парадак дзеразовыя), разнаспоравым першасна травяністым (парадак селагінелавыя) і разнаспоравым травяністым і дробным драўнінным (парадак проталепідадэндравыя). Ад проталепідадэндравых узялі пачатак разнаспоравыя буйныя лепідадэндравыя і буйныя драўнінныя сігілярыевыя (парадак лепідадэндравыя). Ад разнаспоравых дробных драўнінных лепідадэндравых узялі пачатак разнаспоравыя другасна травяністыя палушнікавыя (парадак палушнікавыя).