 2013-12-31
2013-12-31 749
749Пытанні і заданні для самакантролю
1. Дайце характарыстыку псілотападобных – «жывых выкапнёвых» сучаснай флоры.
2. Дайце характарыстыку хвошчападобных.
3. Вызначце асаблівасці цыкла развіцця хвашчу палявога.
1.Назавіце асаблівасці раслін аддзела папарацепадобных.
2.Якія ўмовы развіцця ўласцівы вужоўнікавым папарацям?
3.Ахарактарызуйце цыкл развіцця раўнаспоравых папарацей на прыкладзе шчытоўніка мужчынскага.
4.Апішыце цыкл развіцця сальвініі плывучай як прадстаўніка разнаспоравых папарацей.
5.Якое філагенетычнае значэнне мае разнаспоравасць у папарацей?
НАСЕННЫЯ РАСЛІНЫ
Насенныя расліны характарызуюцца размнажэннем пры дапамозе насення, якое развіваецца з семязачатка. Семязачатак – гэта відазменены мегаспарангій.
У насенных раслін гаметафіты рэдукаваны яшчэ больш, чым у папарацепадобных.
Пры прарастанні мегаспоры ўнутры семязачатка ўтвараецца жаночы гаметафіт (першасны эндасперм) з архегоніямі на мацярынскай расліне ў голанасенных, ці 8 ядзернага зародкавага мешка ў пакрытанасенных.
Моцна рэдукаваны мужчынскі гаметафіт (прарослая пылінка) пераносіцца тым ці іншым чынам на семязачатак. У сваёй вегетатыўнай частцы ён зведзены да ўтварэння галоўным чынам пылковай трубкі. Пылковая трубка праводзіць мужчынскія гаметы, якія страцілі ў большасці насенных раслін рухомасць, непасрэдна да жаночай гаметы (яйцаклеткі), схаванай унутры семязачатка, адбываецца працэс апладнення і развіцця зародка. Гэта забяспечвае незалежнае апладненне ад вады, яго аўтаномнасць. У працэсе развіцця зародка семязачатак ператвараецца ў семя – асноўны фактар рассялення раслін. У пераважнай большасці насенных раслін гэтае ператварэнне семязачатка ў спелае, гатовае да прарастання семя адбываецца на самой мацярынскай расліне.
Зародак звычайна ўтрымлівае ўсе асноўныя органы дарослай расліны: сцябло, ліст, корань. Ён утрымлівае таксама запас спажыўных рэчываў, добра ахаваны насеннай лупінай ад неспрыяльных атмасферных і фізічных уздзеянняў.
Семя аддзяляецца ад мацярынскай расліны і, упаўшы на зямлю, можа доўгі час знаходзіцца ў стане спакою. З надыходам спрыяльныў умоў (цяпла, вільгаці і інш.) семя прарастае. Зародак кранаецца ў рост, выкарыстоўваючы ў пачатковы перыяд свайго самастойнага развіцця запасныя рэчывы семені. Лупіна семені скідаецца, праростак ўкараняецца ў зямлю і расліна пераходзіць да самастойнага жыцця.
Унутранае апладненне, развіццё зародка ўнутры семязачатка і з‘яўляецца новым, надзвычай эфектыўным фактарам рассялення, галоўнымі біялагічнымі перавагамі насенных раслін, даўшымі ім магчымасць лепш прыстасоўвацца да наземных умоў і дасягнуць больш высокага ўзроўню развіцця, чым у папарацей і іншых безнасенных вышэйшых раслін. Калі пры размнажэнні спорамі кожны раз утвараецца велізарная іх колькасць – цэлыя мільёны, дык пры размнажэнні насеннем яго колькасць у многа разоў меншая. Гэта пацвярджае, што семя – больш надзейная адзінка рассялення, чым спора. У семені ўжо ўтрымліваецца, пры тым у вельмі добрай і надзейнай упакоўцы, зародак – маленечкі спарафіт з карэньчыкам, пупышкай і зародкавымі лістамі (семядолямі). У гэтым істотная перавага насенных раслін перад споравымі і разнаспоравымі раслінамі.
Цудоўна прыстасаваўшыся да сучасных умоў жыцця на зямлі, насенныя расліны істотна пацяснілі споравыя расліны і занялі пануючае становішча на сушы.
Насенныя расліны падзяляюцца на два аддзелы: голанасенныя (Рinophyta, або Gymnospermae) і пакрытанасенныя, ці кветкавыя расліны (Аngiospermae, ці Мagnoliophyta).
АДДЗЕЛ ГОЛАНАСЕННЫЯ (PINOPHYTA, АБО GYMNOSPERMAE)
Паходжанне голанасенных раслін
Паходжанню голанасенных раслін папярэднічала некалькі эвалюцыйных падзей, якія адбыліся ў дэвонскім перыядзе (каля 400 млн. гадоў назад): узнікненне разнаспоравасці, камбію, з‘яўленне драўнінных форм.
Гэта вельмі старажытная група вышэйшых раслін. У якасці магчымых продкаў голанасенных разглядаюць старажытнейшых прадстаўнікоў аддзела папарацепадобных. Менавіта сярод іх мы знаходзім разнаспоравыя драўняныя формы з другаснай драўнінай.
Самым старажытным класам аддзела голанасенных былі расліны класа насенныя папараці, а найбольш дасканалымі з‘яўляюцца расліны класа гнетапсіды, якія філагенетычна найбольш блізкія да раслін аддзела пакрытанасенныя.
Амерыканскія батанікі ўказваюць на паходжанне голанасенных ад рыніяфітаў праз трымерафітаў і прагімнаспермаў. Насенныя папараці і кардаіты з голанасенных дасягаюць вялікай складанасці ўжо ў каменнавугальным і пермскім перыядах. Астатнія прадстаўнікі голанасенных былі добра развіты ў мезазоі. Мезазой – эра панавання голанасенных (бенетытавыя, сагаўнікавыя, гінкгавыя).
Голанасенныя – разнаспоравыя насенныя расліны. Семязачаткі (мегаспарангіі з акружаючымі іх інтэгументамі) у голанасенных на мегаспарафілах размешчаны адкрыта, гола. Адсюль і назва аддзела. Яны налічваюць каля 800 відаў.
У цыкле развіцця голанасенных, як і ў большасці споравых, пераважае спарафіт. Гаметафіт моцна рэдукаваны. Пры гэтым жаночы гаметафіт не пакідае сувязі з мацярынскай раслінай, развіваецца ўнутры семязачатка, дзе адбываецца ўтварэнне мегаспоры і яе прарастанне ў жаночы зарастак (першасны эндасперм). Мужчынскі гаметафіт таксама зазнае рэдукцыю аж да страты вегетатыўных (праталіяльных) клетак у некаторых прадстаўнікоў.
Сучасныя голанасенныя – выключна дрэвавыя расліны (дрэвы, хмызнякі, ліяны). Сярод іх няма траў.
Лісты голанасенных сінтэломнага паходжання, эвалюцыя якіх ішла ў двух напрамках. У найбольш старажытных відаў з неразгалінаванымі парасткамі захаваліся працягла жывучыя буйныя лісты з рассечанай ліставой пласцінкай. Са з’яўленнем разгалінавання адбывалася змяншэнне памераў лістоў, што аказалася асабліва важным ва ўмовах халоднага і сухога клімату. У цяперашнія часы большасць голанасенныя адносіцца да хвойных.
Аддзел уключае шэсць класаў: насенныя папараці, ці лігінаптэрыдапсіды (Lyginopteridopsida), сагаўнікавыя (Cycadopsida), бенетытавыя (Bennettitopsida), гінкгавыя (Ginkgoopsida), хвойныя (Pinopsida), гнетавыя,ціабалонканасенныя (Gnetopsida, ці Chlamydospermatopsida).
У некаторых сістэмах класам голанасенных адпавядаюць аддзелы.
КЛАС НАСЕННЫЯ ПАПАРАЦІ (LYGINOPTERIDOPSIDA)
Гэта самая старажытная, цалкам вымерлая група голанасенных, якая жыла з сярэдзіны дэвону да мелавога перыяду (350-285 млн. гадоў назад).
Насенныя папарацці былі дрэвападобнымі, ліянападобнымі, травяністымі раслінамі з вертыкальнымі ці ляжачымі сцёбламі і даволі буйнымі простымі, перыстарассечанымі лістамі, як у сучасных папарацей; у некаторых лісты былі цэльныя.
Не менш разнастайнай была анатамічная будова сцёблаў насенных папарацей. У адных відаў камбій функцыянаваў вельмі слаба, адкладваў толькі некалькі слаёў другаснай ксілемы (у травяністага тыпу сцябла), у другіх раслін мелася моцная драўніна тыпу хвойных, у трэціх – большую частку сцябла займала парэнхіма асяродка, як у сагаўнікавых. Падобная разнастайнасць марфалагічнай і анатамічнай будовы розных раслін у межах аднаго перыяду іх жыцця дазваляе меркаваць, што стварэнне жыццёвых форм і анатамічных структур як гола-, так і пакрытанасенных з самага пачатку магло пайсці рознымі шляхамі, ці маглі ўзнікнуць першасна драўнінныя і першасна травяністыя формы. Таму з гэтай групы насенных папарацей лёгка вывесці ўсе іншыя класы голанасенных раслін.
Вялікую цікавасць уяўляе разнастайнасць у будове семязачаткаў, якая пралівае святло на паходжанне інтэгумента. Самыя старажытныя семязачаткі мелі вельмі прымітыўную будову – нуцэлус (мегаспарангій) з адцягнутым на вяршыні носікам быў акружаны вузкімі, адцягнутымі вонкі лопасцямі інтэгумента. Лопасці зрасталіся паміж сабой і з нуцэлусам толькі ў самай аснове; у кожную лопасць уваходзіў праводзячы пучок. Гэты лопасцевы інтэгумент нагадваў сукупнасць збліжаных і пачаўшых зрастацца вегетатыўных тэломаў, што падмацоўвае пазіцыі сінтэломнай тэорыі паходжання інтэгумента. У большасці семязачаткаў, апісаных з сярэдняга і верхняга карбону, лопасці інтэгументаў цалкам ці амаль цалкам зрасталіся паміж сабой, заставаліся свабоднымі толькі на вяршыні; у кожную лопасць уваходзіла жылка. Паралельна з гэтым адбывалася зрастанне інтэгумента з нуцэлусам. У многіх насенных папарацей асобныя семязачаткі ці групы семязачаткаў размяшчаліся ў акружэнні чашападобнага ўтварэння – купулы. Адны аўтары разглядаюць яе як вырасты ліста (па наяўнасці залозак на яе паверхні), іншыя – як вынік зрастання перыферыйных тэломаў. Купула ўяўляе сабой сукупнасць двойчы ці тройчы дыхатамічна разгалінаваных лопасцей, у кожную з якіх уваходзіць жылка. Лопасці купулы зліваліся толькі ў аснове. У сярэдне- і верхнекарбонавых прадстаўнікоў лопасці купулы з праводзячымі пучкамі зрасталіся на большым сваім працягу ці цалкам. Купулы, у сваю чаргу, маглі зрастацца ў большай ці меншай ступені з інтэгументам семязачатка, утвараючы знешні інтэгумент (купуламент), пранізаны пучкамі. Падобны двайны інтэгумент маюць семязачаткі сагаўнікавых. У больш позніх голанасенных магло адбывацца поўнае зліццё ўнутранага інтэгумента з нуцэлусам амаль да знікнення праводзячых пучкоў. У гэтым выпадку семязачаткі з адным знешнім інтэгументам станавіліся другасна аднапокрыўнымі.
Такім чынам, першыя голанасенныя маглі мець 3 варыянты будовы семязачаткаў – зыходна двухпокрыўных, ці з адным інтэгументам, двухпокрыўныя – з унутраным і знешнім інтэгументамі і другасна аднапокрыўнымі са знешнім інтэгументам. Акрамя таго, у межах гэтых трох варыянтаў семязачаткі мелі радыяльную ці дарсівентральную сіметрыю, ці лік варыянтаў будовы семязачаткаў павялічваўся да 6-ці. Меліся яшчэ дадатковыя варыянты семязачаткаў. Будова семязачаткаў аказалася вельмі стабільнай і характэрнай для многіх класаў голанасенных. Разнастайнасць семязачаткаў дазволіла ўстанавіць пераемнасць рэпрадуктыўных органаў пакрытанасенных ад голанасенных раслін.
Семязачаткі ў адных відаў размяшчаліся на звычайных лістах, у другіх – на спецыяльных лістах - мегаспарафілах, у трэціх – на восевых структурах-мегаспарангіяфорах. Цікавая асаблівасць насення насенных папарацей заключаецца ў адсутнасці зародкаў; верагодна, насенне ападала раней, чым адбывалася іх поўнае фарміраванне, як у сагаўнікаў. Мікраспарангіі часта аб‘ядноўваліся ў сінангіі, якія, як правіла, размяшчаліся на спецыялізаваных перках вегетатыўных лістоў, ці меў месца дымарфізм частак ліста. Форма спараносных частак ліста была самая разнастайная. У мікраспарангіях некаторых відаў знойдзены мужчынскія гаметафіты; у адных відаў лік праталіяльных клетак даходзіў да 30-ці, а ў некаторых іх было ўсяго толькі 2.
Розныя органы насенных папарацей апісаны пад рознымі назвамі: лігінаптэрыс (Lyginopteris), кросатэка (Crossotheca), археасперма (Archaeosperma), трыгонакарпус (Trigonocarpus) і іншыя.
Прадстаўнікі: калімататэка(Calymmatotheca, малюнак 43), медулоза (Medullosa), кейтонія (Caytonia).
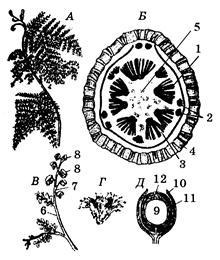

КЛАС САГАЎНІКАВЫЯ (CYCADOPSIDA)
У цяперашні час сагаўнікавыя прадстаўлены адным парадкам (Cycadales), адной сям’ёй (Cycadaceae), 9-10 родамі і 100-120 відамі ў трапічных і субтрапічных абласцях.
Сагаўнікі з‘яўляюцца рэшткамі некалі шырока распаўсюджанай групы раслін. Выкапнёвыя рэшткі сагаўнікаў (лісты і насенне) сустракаюцца ў адкладаннях ранняпермскага перыяду. Па апошніх дадзеных сагаўнікавыя з‘явіліся ў познім карбоне, дасягнулі росквіту ў юрскім і мелавым перыядах і звязаны ў сваім паходжанні з насеннымі папарацямі, якіх яны нагадваюць многімі прыкметамі сваёй знешняй і ўнутранай арганізацыі. Сучасныя
сагаўнікі – дрэвы з калонападобным сцяблом вышынёй да 6-20м ці клубнепадобным, невысокім, схаваным у зямлі (малюнак 44), вельмі нямногія эпіфіты на ствалах дрэў.
Родсагаўнік (Cycas) налічвае 16 відаў. Распаўсюджаны ад Усходняй Азіі да Паўночна-Усходняй Аўстраліі і на востраве Мадагаскар. Цяпер жывучыя сагаўнікі з’яўляюцца рэшткамі некалі шырока распаўсюджанай групы раслін. Выкапнёвыя рэшткі сагаўнікаў сустракаюцца ў адкладаннях трыясу і юры. Тады яны былі панавальнымі сярод іншых наземных раслін.


У заміімнагабрачнай (Zamia polygama), якая расце на Кубе, сцябло вельмі маленькае – 3см увышыню і 2см у дыяметры.
Лісты сагаўнікаў сцябловага паходжання, простаперыстыя, як у пальмаў, радзей двойчы- і тройчыперыстыя, да 2м удаўжыню, цвёрдыя, вечназялёныя, у маладым стане ўліткападобна загорнутыя, як у папарацей.
Сцябло мае асяродак з крухмалам, за ім ідзе вузкае кальцо слаба развітай рыхлай драўніны, затым ідзе камбій, слой флаэмы і тоўстая кара. У кары маюцца слізевыя хады са сліззю (падабенства з мараціевымі папарацямі). Такі тып будовы сцябла называюць манаксільным (ад грэч. manos – рыхлы і xylos – драўніна).
Корань сагаўнікаў доўгі, разгалінаваны. Для каранёвай сістэмы сагаўнікаў характэрным з’яўляецца наяўнасць бакавых паверхневых каранёў, якія растуць уверх. У паверхні зямлі ці крыху вышэй яны моцна разгаліноўваюцца, утвараюць каралападобныя ці клубнепадобныя скапленні, якія размяшчаюцца каля асновы ствала. У пратадерме гэтых каранёў былі выяўлены сіне-зялёныя водарасці (Nostok punctiforme, Anabaena cycadae, і інш.), азотфіксавальныя бактэрыі (Bacterium radicicola i Azotobacter sp.) і нарэшце гіфы слаба патагенных грыбоў, утвараючых фікаміцетную эндатрофную мікарызу. Роля гэтых каранёў, верагодна, заключаецца ў забяспячэнні расліны азоцістымі злучэннямі.
Спарангіяфоры сабраны ў стробілы. Сагаўнікі – двухдомныя расліны. На верхавіне сцябла мужчынскага экзэмпляра знаходзяцца мікрастробілы даўжынёй ад 1,5-2,0 да 50-70см. На восі мікрастробілы размешчаны мікраспарангіяфоры(малюнак 45), якія адрозніваюцца ад вегетатыўных лістоў па форме (трохвугольнай, шматвугольнай, грыбападобнай і інш.). На ніжнім баку мікраспарангіяфораў фарміруюцца мікрасорусы з 2-4 мікраспарангіяў. У мікраспарангіях развіваецца вялікая колькасць эліпсаідальных дробных спор. Мікраспоры без паветраных мяшкоў. Мікраспора пачынае дзяліцца ўнутры мікраспарангія, фарміруе пад сваёй абалонкай мужчынскі гаметафіт, які складаецца з трох ці чатырох клетак (праталіяльнай, гаўстарыяльнай і стэрыльнай), разносіцца ветрам.
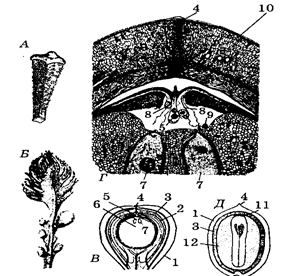

У большасці сагаўнікавых ёсць шышкі (стробілы). Яны адсутнічаюць толькі ў сагаўніка. У сагаўніка семязачаткі размяшчаюцца на жоўта-аранжавых перыстых мегаспарафілах у ніжняй іх частцы, на разгалінаваннях, у колькасці 6 па 3 з кожнага боку мегаспарафіла. Семязачаткі дасягаюць 5-6 см удаўжыню.
Развіццё семязачатка адбываецца наступным чынам. На мегаспарафіле ўзнікае нуцэлус у выглядзе невялікага бугарка з аднолькавых клетак. Па меры яго росту ад асновы нуцэлуса вырастае інтэгумент. Ен моцны, трохслойны. Знешні і ўнутраны слаі мясістыя з праводзячымі пучкамі (васкулярызаваныя), а паміж імі – сярэдні камяністы слой. Інтэгумент не поўнасцю змыкаецца ўверсе і ўтварае мікропілі – пылкаўваход. У верхняй частцы нуцэлуса ў выніку аслізнення клетак узнікае пылковая камера са сліззю, кропелька якой выступае праз мікропіле. Паходжанне інтэгумента тлумачыцца па-рознаму.
Сінангіяльная гіпотэза паходжання інтэгумента семязачатка наступная. Апошні ўзнік з сінангія (групы спарангіяў), у якім унутраны спарангій застаўся фертыльным (прадуцыруе споры), а перыферычныя спарангіі падвергліся стэрылізацыі і ўтварылі інтэгумент.
Па тэломнай гіпотэзе інтэгумент ўяўляе сабой вынік зрастання перыферычных зыходна вегетатыўных тэломаў вакол аднаго спараноснага тэлома.
У верхняй частцы нуцэлуса адасабляецца адна буйная клетка – мацярынская клетка мегаспор. Яна рэдукцыйна дзеліцца з утварэннем 4-х гаплоідных мегаспор. Яны размяшчаюцца ланцужком адна над другой па вертыкалі. Тры верхнія мегаспоры адміраюць, а ніжняя прарастае і ўтварае жаночы зарастак (гаметафіт) – першасны эндасперм. Зарастак, такім чынам, знаходзіцца ўнутры семязачатка. На жаночым зарастку фарміруецца два архегоніі. Архегоній складаецца з буйной яйцаклеткі, рана адміраючай брушной канальцавай клеткі і дзвюх шыйкавых клетак.
Над архегоніямі перыферычныя клеткі зарастка разрастаюцца і фарміруюць унутраную поласць, называемую археганіяльнай камерай. Паступова дно пылковай камеры разбураецца і яна (пылковая камера) зліваецца з археганіяльнай камерай.
Прарослая мікраспора сагаўніка трапляе на семязачатак і прыклейваецца да кропелькі слізі, якая выступае праз мікропіле семязачатка. Па меры падсыхання вадкасці пылінка ўцягваецца ў поласць, у так званую пылковую камеру. Тут пылінка пачынае прарастаць з утварэннем гаўсторыі, якая прарастае ў тканку нуцэлуса, паглынае спажыўныя рэчывы і вільгаць. Сперматагенная клетка дзеліцца і ўтварае 2 сперматазоіды памерам да 3-3,5 мм са шматлікімі (некалькі тысяч) жгуцікамі па спіралі. Гэта самыя буйныя сперматазоіды ў раслінным свеце. Унутры пылінкі ствараецца высокі тургарны ціск за кошт паглынання ёю вільгаці з клетак нуцэлуса. Гэта прыводзіць да разрыву гаўстарыяльнай трубкі. Пад вялікім ціскам слізь са сперматазоідамі выкідаецца на дно археганіяльнай камеры. Канальцавыя клеткі архегонія аслізняюцца. Гэта забяспечвае доступ сперматазоідаў да яйцаклеткі. Перыяд ад апылення да апладнення працягваецца 6-7 месяцаў. Зігота фарміруе ў верхняй частцы падвесак, ці суспензар, які прасоўвае зародак у тканку жаночага гаметафіта, багатага спажыўнымі рэчывамі. Зародак заканчвае сваё фарміраванне ў апалым насенні на працягу 1-2-х гадоў. Ён складаецца з карэньчыка, сцяблінкі, пупышкі і ад 1 да 6, часцей 2-х семядолей. Семя без перыяду спакою прарастае. Гэта разглядаецца як вельмі прымітыўная прыкмета ў сагаўнікаў.
Цыкл развіцця прадстаўніка сагаўнікавых заміі (Zamia) прыведзены на малюнку 46.
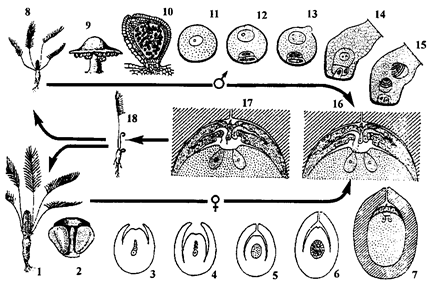

Неабходна адзначыць, што нуцэлус гамалагічны мегаспарангію папарацепадобных, з’яўляецца новым утварэннем у насенных раслін.
Прыкметы старажытнасці сагаўнікавых праяўляюцца ў наступным: 1) макрафілія; 2) слабае развіццё ў сцябле драўніны і наяўнасць моцнага асяродка; 3) апладненне з дапамогай сперматазоідаў; 4) прарастанне насення без перыяду спакою.
Сагаўнікі знаходзяць практычнае прымяненне. З асяродка атрымліваюць сагавую крупу. Многія сагаўнікі – прыгожыя дэкаратыўныя расліны.
КЛАС БЕНЕТЫТАВЫЯ (BENNETTITOPSIDA)
Выкапнёвыя расліны жылі на Зямлі ад трыясавага да мелавога перыяду мезазою (230-70 млн. гадоў назад).
Выміранне бенетытавых супала з узнікненнем і росквітам кветкавых раслін. Гэта былі невялікія дрэвы аблічча сагаўнікаў ці папарацей. Іх сцёблы былі простымі ці слаба разгалінаванымі, па вышыні да 2м ці прыземістымі клубнепадобна й формы дыяметрам да 60см. Лісты ў большасці былі складанаперыстымі, радзей простымі, цэльнымі. Па многіх асаблівасцях знешняй і внутранай будове яны былі падобны на сагаўнікавыя.
Асаблівасць бенетытавых у тым, што некаторыя з іх мелі орган размнажэння – двухполую пазушную шышку, стробіл (малюнак 47). Звонку шышка была адзета верхавіннымі лістамі, размешчанымі па спіралі. Далей па восі шышкі размяшчаліся кругамі перыстыя (радзей цэльныя) мікраспарангіяфоры з мікраспарангіямі. Шматлікія мікраспоры былі без паветраных мяшкоў. У верхняй частцы стробіла, заканчваючы вось, размяшчаліся мегаспарангіяфоры ў выглядзе доўгіх цыліндрычных ножак з семязачаткамі, на вяршынях якіх былі моцна выцягнутыя мікрапілярныя трубачкі. Яны чаргаваліся са стэрыльнымі органамі, якія на верхавіне шматвугольна пашыраліся ў шчыткападобную пласцінку, ахопліваючы з бакоў семязачатак і ахоўваючы яго. Таму бенетытаў называюць ”паўпакрытанасеннымі” раслінамі. Усе семязачаткі мелі пылковую камеру, у сувязі з чым можна меркаваць, што апладненне ў іх адбывалася з дапамогай сперматазоідаў, як у сагаўнікавых.
У некаторых бенетытавых былі раздзельнаполыя (другасна) шышкі.


Бенетыты валодалі ўнікальным сярод голанасенных раслін насеннем, якое не мела эндасперма. Іх спажыўныя рэчывы змяшчаліся ў двух буйных семядолях, што лічыцца прагрэсіўнай асаблівасцю.
Бенетыты паходзяць, верагодна, ад птэрыдаспермавых. Некаторыя лічаць бенетытавых магчымымі продкамі пакрытанасенных. Пасля адкрыцця бенетытаў ix шышку (стробіл) сталі разглядаць як прататып кветкі пакрытанасенных раслін. Англійскія палеабатанікі Э. Арбер і Дж. Паркін стварылі страбілярную гіпотэзу паходжання кветкі раслін. На самой справе, двухполая шышка бенетытаў знешне падобна на кветку магноліі, але пакрытанасеннасць у яе іншай прыроды, чым у кветкі пакрытанасенных раслін.
КЛАС ГІНКГАВЫЯ (GINKGOOPSIDA)
Клас гінкгавыя ўключае парадак Ginkgoales. У цяперашні час прадстаўлены адзіным відам – гінкга двухлопасцевы (Ginkgo bіloba). У дзікім стане гэты від сустракаецца толькі ў гарах Усходняга Кітая, але з даўніх часоў шырока культывуецца і ў раёнах з умераным кліматам.
Гінкга – “жывы выкапнёвы” прадстаўнік класа. У мезазоі гінкгавыя былі шырока распаўсюджаны на зямлі (Еўразія, Паўночная Амерыка, Паўднёвае паўшар’е). Выміранне гінкгавых пачалося з мелавога перыяду.
Гінкга двухлопасцевы – лістападнае дрэва больш 40 м увышыню і больш 3 м у дыяметры. Мае парасткі двух тыпаў – падоўжаныя (аўксібласты) і пакарочаныя (брахібласты). На падоўжаных парастках лісты размешчаны адзіночна, на пакарочаных – сабраны пучкамі па 5-7. Лісты чаранковыя, веерападобныя, двухлопасцевыя, з дыхатамічным жылкаваннем. Вясной і летам лісты светла-зялёныя, увосень – жаўтаватыя, кармінава-чырвоныя.
Анатамічная будова сцябла тыповая для хвойных: слабаразвіты асяродак, другасная драўніна пікнаксільная (ад грэч. pyknos – шчыльны і xylos – драўніна) з трахеід i асяродкавых прамянёў, камбій, кара.
Гінкга – расліна двухдомная (малюнак 48).
Мужчынскае “суквецце” мікрастробіл мае выгляд каташка. Каташкі размешчаны на верхавіне скарочанага парастка. На восі каташка размешчаны мікраспарангіяфоры. Кожны мікраспарангіяфор нясе звычайна 2, радзей 3-7 мікраспарангіяў. Мікраспоры без паветраных мяшкоў. Мікраспоры прарастаюць у зарастак яшчэ ўнутры мікраспарангія.
Прарастанне мікраспоры гінкга, працэс утварэння мужчынскага гаметафіта, апладненне, фарміраванне зародка і насення ў цэлым падобна на сагаўнікавыя.
Семязачаткі ў колькасці двух развіваюцца на верхавіне скарочанага парастка. Кожны семязачатак акружаны ў аснове кальцавым валікам (каўнерыкам), які з‘яўляецца рэдукаваным мегаспарафілам. Будова семязачатка такая, як у сагаўніка. Аплодненая яйцаклетка дзеліцца, утварае зародак з карэньчыкам, сцяблінкай і семядолямі; маецца і падвесак. Семязачатак становіцца насеннем. Насенне адзета тоўстай абалонкай з інтэгумента. Знешняя частка абалонкі становіцца сакавітай, пад ёю размешчана цвёрдая камяністая абалонка і ўнутраны тонкі плевачны слой. Зародак у насенні акружаны эндаспермам. Насенне прарастае без бачнага перыяду спакою, што адносіцца да ліку прымітыўных прыкмет.


Рысамі прымітыўнай арганізацыі гінкга з’яўляюцца: дыхатамічнае жылкаванне лістоў; прысутнасць пылковай камеры, запоўненай вадкасцю; рухомыя мужчынскія гаметы – сперматазоіды; прарастанне насення без перыяду спакою.
Насенне гінкга выкарыстоўваецца ў медыцыне і ўжываецца ў ежу. У якасці дэкаратыўнай расліны гінкга выкарыстоўваецца для азелянення паўднёвых гарадоў. Вырошчваецца і ў Беларусі.








