2018-03-09
2018-03-09 149
1491. В коде отсутствуют знаки препинания, поэтому рамка считывания должна быть правильно установленной в начале прочтения молекулы мРНК, а затем должна двигаться последовательно от одного триплета к следующему. Если исходная рамка считывания «сбита» на один или два нуклеотида или же рибосома случайно пропустила один нуклеотид в мРНК, все последующие кодоны выйдут из правильной рамки и это приведет к образованию полипептида с искаженной аминокислотной последовательностью;
2. Из 64 возможных триплетов три (UAG, UAA, UGA) не кодируют ни одну из аминокислот _ это нонсенс-кодоны, которые сигнализируют об окончании синтеза полипептида. Кодон AUG представляет собой инициирующий кодон и у прокариот, и у эукариот; кроме того, во внутренних положениях полипептидной цепи он кодирует метионин.
3. Кодовые «слова» аминокислот одинаковы у всех изученных организмов, включая человека, бактерии, растения, земноводных и вирусы. Создается впечатление, что все виды растений и животных имели общего эволюционного предшественника с одним генетическим кодом, полностью сохранившимся на протяжении всей биологической эволюции. Таким образом, код практически универсален. Однако появились факты, которые говорят о том, что в некоторых случаях при синтезе белка митохондриями в присутствии рибосом, тРНК и мРНК митохондриального происхождения ряд аминокислотных кодонов используется не в соответствии с их значением по стандартному кодовому «словарю».
ТЕРМИНАЦИЯ
Терминация – окончание биосинтеза полипептида - наступает после присоединения рибосомой последней аминокислоты. О терминации сигнализирует один из трех терминирующих кодонов мРНК (UAA, UAG, UGA). Их называют бессмысленными триплетами (нонсенс-триплетами). При этом начинают действовать три терминирующих фактора (факторы освобождения, рилизинг-факторы) - белки R1, R2, S. Они вызывают: 1) гидролитическое отщепление полипептида от конечной тРНК и его высвобождение; 2) отделение от Р-участка последней, теперь уже пустой тРНК; 3) диссоциацию 70S-рибосомы на 30S- и 50S-субчастицы, готовые к синтезу новой полипептидной цепи. Фактор R1 узнает кодоны UAA или UAG. Второй фактор освобождения (R2) узнает UAA или UGA. Связывание одного из факторов освобождения с терминирующим кодоном в А-участке активирует пептидилтрансферазу, и она гидролизует связь между полипептидом и тРНК в Р-участке. Происходит изменение специфичности пептидилтрансферазы таким образом, что акцептором активированного пептидильного остатка становится Н2O, а не аминогруппа.
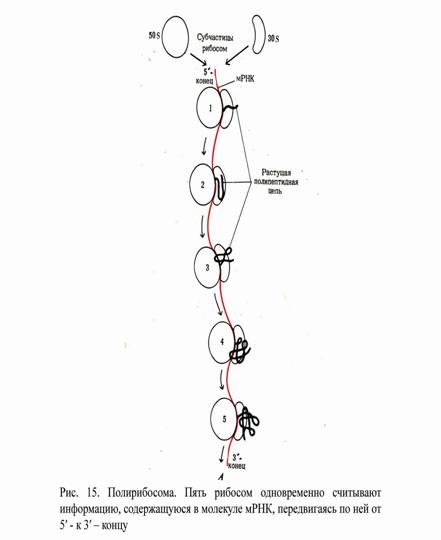
Часто рибосомы собраны в группы, состоящие из нескольких десятков рибосом. Это полирибосомы, или полисомы. Они были изучены с помощью электронного микроскопа, а также химическим путем.
Под действием рибонуклеазы полисомы разобщаются на отдельные рибосомы. Это указывает на то, что они удерживаются с помощью цепи РНК. Итак, мРНК одновременно транслируется многими рибосомами, расположенными довольно близко друг к другу. Такая одновременная трансляция одной мРНК многими рибосомами значительно увеличивает эффективность использования матрицы (рис.15).
Процессы транскрипции и трансляции в бактериях очень тесно сопряжены. Рибосомы могут начать транслировать мРНК, когда та еще продолжает синтезироваться ДНК-зависимой РНК-полимеразой (рис.16). Другая особенность белкового синтеза в бактериях заключается в том, что время жизни молекул мРНК очень мало, всего несколько минут: они быстро разрушаются нуклеазами. Чтобы синтез белка поддерживался на одном уровне, мРНК для данного белка или для группы белков должна синтезироваться постоянно и использоваться с максимальной эффективностью. Короткое время жизни мРНК у прокариот позволяет быстро выключать синтез белка, который больше не нужен.