 2018-02-13
2018-02-13 984
984В проводящей системе сердца, а также в самой сердечной мышце могут образовываться замкнутые пути, по которым циркулирует волна возбуждения. Модельно это явление можно представить последовательностью прохождения двух волн возбуждения в гипотетическом кольце. Если кольцо однородно по рефрактерности, то две волны возбуждения, идущие по кольцу от источника возбуждения А, аннигилируют в точке В. Если в кольце активной среды имеется участок CD, период рефрактерности элементов которого R2больше, чем период рефрактерности остальной среды Н^рис. 6.6), то в этом случае может возникнуть циркуляция возбуждения в нем. Это произойдет, если в точке внешнее воздействие создает подряд два возбуждения. Причем вторая волна возникает через время, меньшее периода рефрактерности участка СД: T<R2. Тогда волна II может дойти до участка СД к моменту времени, когда он еще остается рефрактерным, и гасится. Остается одна волна I. Если она дойдет до участка СД через время, за которое он успеет прийти в состояние покоя, волна I пройдет дальше и в кольце так и будет продолжаться незатухающий процесс - циркуляция возбуждения.
Условия возникновения циркуляции:
1) время между посылкой двух импульсов возбуждения Т должно быть меньше периода рефрактерности R2: Т < R2V.
2) длина окружности кольца I = 2тсг должна быть больше длины волны возбуждения:
Длина циркулирующей волны в путях проведения при V = 3 м/с и R = 0,3 с может составлять величину около 1 м, что анатомически исключает ее появление в этих путях. Однако в узлах медленного проведения возбуждения (V = 0,01 м/с) ё может иметь порядок нескольких миллиметров и в этом случае механизм циркуляции волны возбуждения может быть реализован в сердце. Такая рециркуляция может наблюдаться в области атриовентрикулярного узла и в зонах с замедленным проведением автоволны.
20. Различные типы взаимодействий в макромолекулах.
Первичная структура или последовательность звеньев в полимерной цепи(например, аминокислоты в цепи белка, нуклеотиды в цепи нуклеиновых кислот). Определяется физическими валентными связами кроме того существует слабые не валентные силы проводящие притяжение на больших расстояниях и отталкивание на малых расстояниях.
1. Силы Ван-дер-Ваальсовые- эти силы играют важную роль в образовании конденсированных жидких и твердых состояний. Ими определяются взаимодействия газов и возникающие отклонения законов идеальных газов.  где R- газовая постоянная, a, b - константы притяжения и отталкивания; V -объем.
где R- газовая постоянная, a, b - константы притяжения и отталкивания; V -объем.
Различают ориентационные и дисперсионные типы сил Ван-дер-Ваальса.
2. Водородная связь – она осуществляется между атомом водорода одной молекулы и электроотрицательным атомом кислорода (О), азота (N), фтора (F), или хлора (Cl), принадлежащего другой молекуле. Энергию водородной связи вносят вклад: энергия отталкивания, энергия дисперсного взаимодействия и энергия делокализации 2-х электронов связи (А-Н) и неподеленной пары электронов другого атома. Uвод=Uотталк+Uдисп+Uэл. стат+Uделок
3. Электростатическое взаимодействия. Помимо того что вносят вклад энергию водородной связи, эти взаимодействия играют роль стабилизация биоструктур, частично заряженных атомов описывается электростатическим потенциалом вид которого задается уравнением:
 где
где 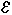 - проницаемость, q-величина заряда на атомов i и j, R- расстояние между атомами.
- проницаемость, q-величина заряда на атомов i и j, R- расстояние между атомами.
Структура и функционирование биологических мембран.
Первая модель строения биологических мембран была предложена в 1902 г. Было замечено, что через мембраны лучше всего проникают вещества, хорошо растворимые в липидах, и наосновании этого было сделано предположение, что биологические мембраны состоят из тонкого слоя фосфолипидов. На самом деле, на поверхности раздела полярной и неполярной среды (например, воды и воздуха) молекулы фосфолипидов образуют мономолекулярный (одномолекулярный) слой. Их полярные "головы" погружены в полярную среду, а неполярные "хвосты" ориентированы в сторону неполярной среды. Поэтому и можно было предположить, что биологические мембраны построены из монослоя липидов. В 1925 г. Гортер и Грендел показали, что площадь монослоя липидов, экстрагированных из мембран эритроцитов, в два раза больше суммарной площади эритроцитов. Гортер и Грендел экстрагировали липиды из гемолизированных эритроцитов ацетоном, затем выпаривали раствор на поверхности воды и измеряли площадь образовавшейся мономолекулярной пленки липидов. Однако мембрана - это не только липидный бислой. Имелись экспериментальные данные, которые свидетельствовали о том, что биологическая мембрана состоит и из белковых молекул. Например, при измерении поверхностного натяжения клеточных мембран было обнаружено, что измеренные значения коэффициента поверхностного натяжения значительно ближе к коэффициенту поверхностного натяжения на границе раздела белок-вода (около 10 4 Н/м), нежели на границе раздела липид-вода (около 10~2 Н/м). Эти противоречия экспериментальным результатам были устранены Даниелли и Девсоном, предложившими в 1935 г. так называемую бутербродную модель строения биологических мембран, которая с некоторыми несущественными изменениями продержалась в мембранологии в течение почти 40 лет. Согласно этой модели мембрана - трехслойная. Она образована двумя расположенными по краям слоями белковых молекул с липидным бислоем посередине; образуется нечто вроде бутерброда: липиды, наподобие масла, между двумя "ломтями" белка. Наибольшие успехи в раскрытии особенностей строения биологических мембран были достигнуты в электронно-микроскопических исследованиях. Как известно, световой микроскоп не позволяет рассмотреть детали объекта, меньшие примерно половины длины световой волны (около 200 нм). В световом микроскопе можно разглядеть отдельные клетки, однако он совершенно непригоден для изучения биологических мембран, толщина которых в 20 раз меньше предела разрешения светового микроскопа. Разрешающая способность микроскопа ограничена явлением дифракции. Поэтому, чем меньше длина волны по сравнению с деталями исследуемого объекта, тем меньше искажения. Предел разрешения пропорционален длине волны. В электронном микроскопе вместо светового пучка на исследуемый объект направляется пучок электронов, разогнанных до больших скоростей. Известно, что электронам с высокими скоростями тоже присущи волновые свойства, в том числе явление дифракции. Однако при достаточно больших скоростях, согласно формуле де Бройля, длина волны мала и соответственно мал предел разрешения. Так, если электроны ускоряются электрическим полем с напряжением 105 В, их скорость достигает 10е м/с, длина волны уменьшается и предел разрешения составляет порядка 0,1 нм, что позволяет рассмотреть отдельные детали строения биологических мембран.
Было обнаружено, что имеются белковые молекулы, погруженные в липидный бислой и даже прошивающие его насквозь. Это привело к существенному изменению представлений о строении мембраны.








