2020-05-25
2020-05-25 646
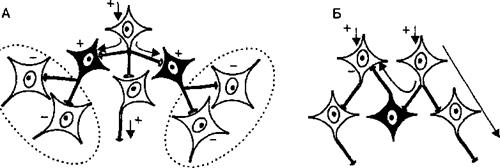
646А — нервная цепочка, Б — дивергенция возбуждения в нервной сети, В — конвергенция возбуждения, Г — реверберация возбуждения в кольцевой цепи Лоренто де Но. Стрелки показывают направление распространения возбуждения.
передается от одного нейрона к другому. Нервная цепь может содержать всего два нейрона с одним синапсом между ними, соответственно рефлекс, реализуемый через такой нервный центр, носит название моносинап- тического. С увеличением числа нейронов и синапсов между ними время рефлекторной реакции на раздражитель увеличивается, поскольку каждый синапс имеет синаптическую задержку проведения. Такие рефлексы называются полисинаптическими. Сигналы по нервным цепям распространяются в одну сторону — от афферентного входа к эфферентному выходу — благодаря одностороннему проведению в синапсах.
Нейроны нервного центра за счет структурно-функциональных связей (ветвления отростков и установления множества синапсов между разными клетками) объединяются в нервные сети. Связи между нервными клетками являются генетически детерминированными.
Различают 3 основных типа нервных сетей: иерархические, локальные и дивергентные с одним входом. Иерархические сети обеспечивают постепенное включение нейронных структур более высокого уровня. В тех случаях, когда афферентная информация поступает к увеличивающемуся числу нейронов, принято говорить о дивергенции возбуждения (рис. 3.9, Б). Если, напротив, от многих возбужденных клеток информация (возбуждение) сходится к меньшему числу нейронов, то такой принцип распространения сигналов называется конвергенцией (рис. 3.9, В). Так, например, конвергенция в эффекторном пути двигательных спинальных рефлексов ведет к тому, что малое число двигательных клеток (мотонейронов) спинного мозга получает импульсы возбуждения от различных эфферентных путей многих рефлекторных дуг. Изучая этот механизм на уровне спинного мозга, Ч. Шеррингтон сформулировал принцип общего конечного пути, согласно которому мотонейроны спинного мозга являются общим конечным путем многочисленных рефлексов (мотонейроны, управляющие сгибателями правой руки, участвуют в многочисленных двигательных рефлекторных реакциях — почесывании, жестикуляции при речи, переносе пищи в рот и т. п.). Соответственно на уровне многочисленных синапсов конвергентных путей возникает конкуренция за общий конечный путь. Однако конвергенция существует и для афферентной информации, когда на ограниченном числе нейронов конвергируют сигналы, приходящие от разных сенсорных систем (слуха, обоняния, зрения и т. п.), что позволяет формировать в нервных центрах полноценный образ воспринимаемого объекта.
Нервные сети обеспечивают реализацию принципа субординации, когда деятельность ниже расположенных нейронных структур подчинена выше расположенным. «Выше» и «ниже» здесь означает уровень расположения в отделах мозга, например, нейроны спинного мозга подчинены влияниям со стороны головного мозга.
Локальные сети нервных центров содержат нейроны с короткими аксонами, осуществляющие взаимосвязи в пределах одного уровня. Они обеспечивают задержку информации в пределах этого уровня, что играет роль в механизмах памяти. Примером такой локальной сети являются кольцевые нейронные цепочки Лоренто де Но, возбуждение в которых циркулирует по замкнутому кругу (рис. 3.9, Г). Возврат возбуждения к «первому»нейро- ну кольцевой цепи получил название реверберации возбуждения. Локальные сети обеспечивают надежность нервной регуляции за счет дублирования элементов, так как многие нейроны локальных сетей имеют одинаковые синаптические связи и функционируют попеременно, т. е. являются взаимозаменяемыми.
Дивергентные сети с одним входом представляют собой нейронные ансамбли, в которых один нейрон образует выходные связи с большим количеством других клеток разных иерархических уровней и, главное, разных нервных центров. Максимально выраженная дивергенция связей разных нервных центров свидетельствует о том, что эти нервные сети не являются специфическими для реализации определенных рефлексов, а обеспечивают интеграцию разных рефлекторных актов и общее состояние активности многочисленных нейронов разных отделов мозга.
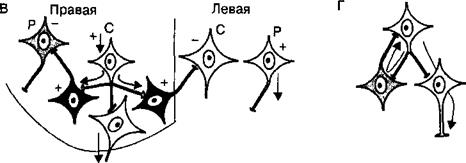
Поскольку в нервных сетях включено огромное число интернейронов, они могут, передавая нервные импульсы другим нейронам центра, реализовывать как их возбуждение, так и торможение. Различают следующие типы тормозных процессов в нейронных сетях:
• реципрокное торможение (рис. 3.10, В), т. е. процесс, основанный на том, что сигналы по одним и тем же афферентным путям обеспечивают возбуждение одной группы нейронов, а через вставочные тормозные клетки вызывают торможение другой группы нейронов. Этот тип торможения называют иначе сопряженным. Проявляется реципрокное торможение, например, на уровне мотонейронов спинного мозга, иннервирующих мышцы-антагонисты (сгибатели-разгибатели) конечностей. Передача возбуждения к мотонейронам мышц-сгибателей одновременно через вставочные тормозные нейроны тормозит мотонейроны мышц-разгибателей, благодаря чему только и возможно осуществление сгибательного рефлекса;
• возвратное торможение (рис. 3.10, Г), т. е. процесс торможения эфферентных нейронов сигналами, возвращающимися к ним по коллатералям аксонов через вставочные тормозные клетки. Возбуждение от одних нейронов поступает через ответвления на аксонах к вставочным тормозным нейронам, а аксоны этих клеток в свою очередь образуют тормозные синапсы на телах первых нейронов. Примером является торможение Реншоу в спинном мозге, где импульсы от двигательных нейронов посылаются по аксонам не только к скелетной мышце, но поступают через аксонные коллатерали к тормозным интернейронам Реншоу, а от них назад к двигательным нейронам, приводя к их торможению. Таким образом осуществляется короткая отрицательная обратная связь, не позволяющая возникнуть избыточному возбуждению мотонейронов спинного мозга;
• латеральное торможение (рис. 3.10, Б), т. е. процесс торможения