 2020-05-25
2020-05-25 1715
1715Генерализованные специализированные эффекты гуморальной регуляции осуществляются с помощью особых химических регуляторов внутренней среды — гормонов. Гормонами называют химические вещества, образующиеся и выделяющиеся специализированными эндокринными клетками (например, р-клетки островков Лангерганса секретируют гормон инсулин), тканями и органами (щитовидной железой, надпочечниками и др.) во внутреннюю среду для регуляции обмена веществ и физиологических функций организма, гуморального обеспечения координации и интеграции процессов жизнедеятельности. Гормоны отличают от других биологически активных веществ, например метаболитов и медиаторов, по двум основным критериям: 1) гормоны образуются специализированными эндокринными клетками, 2) гормоны оказывают свое влияние через внутреннюю среду на удаленные от секретирующей их ткани клетки-мишени, т. е. обладают дистантным действием.
Гормоны являются чрезвычайно высокоактивными химическими соединениями, их концентрации в крови выражаются в нанограммах и даже в пикограммах. Согласно расчетам, 1 г адреналина способен активировать 100 миллионов сердец лягушки, 1 г фолликулина может вызвать течку у 10 миллионов кастрированных мышей, 1 г инсулина — снизить уровень глюкозы в крови у 125 тысяч кроликов. Гормоны оказывают выраженное положительное влияние на эмоциональную сферу, интеллектуальную и физическую активность, выносливость организма, половое поведение.
Эндокринные клетки, образующие гормоны, получили свое название благодаря наличию у них специализированной функции внутренней секреции (инкреции), т. е. активного выведения образовавшихся информационных молекул — гормонов — во внутреннюю среду. Эти специализированные клетки образуют эндокринную систему, т. е. функциональное объединение всех инкреторных клеток организма, осуществляющее гормональную регуляцию.
Гормональная регуляция, как и любая система регуляции, имеет аппарат управления, каналы прямой и обратной передачи информации, сигналы, которыми информация передается, исполнительные органы или объекты управления. Эти элементы системы названы звеньями и составляют структурно-функциональную организацию системы гормональной регуляции.
3.4.1. Общая характеристика звеньев гормональной системы регуляции
Звенья управления. Деятельность эндокринных клеток управляется (регулируется) нервными центрами и гормонами других эндокринных образований, т. е. нейроэндокринным путем. Для некоторых эндокринных клеток основным путем управления является местная саморегуляция за счет механизма обратной связи (например, секреция сахаррегулирующих гормонов* островками Лангерганса регулируется уровнем глюкозы в крови, секреция кальцийрегулирующих гормонов — паратирина и кальцитонина — уровнем кальция в крови), а звенья нейро гуморального управления обеспечивают лишь усиление или ослабление эффекта местной саморегуляции.
Нервная система (рис. 3.12) осуществляет регуляцию деятельности эндокринных клеток двумя путями. Первый из них реализуется структурами центральной нервной системы, непосредственно передающими нервные импульсы к эндокринным структурам, синтезирующим и секретирующим гормоны. Этот путь управления получил название нервного, или парагипо- физарного, т. е. реализуемого мимо гипофиза. Так регулируется деятельность практически всех эндокринных клеток. Второй путь управления эндокринными клетками нервная система реализует через гипофиз, обозначаемый в этом случае как гуморальное звено управления; этот путь регуляции получил название гипофизарного. Таким путем регулируется деятель-
Цереброгландулярный, или парагипофизарный, путь регуляции
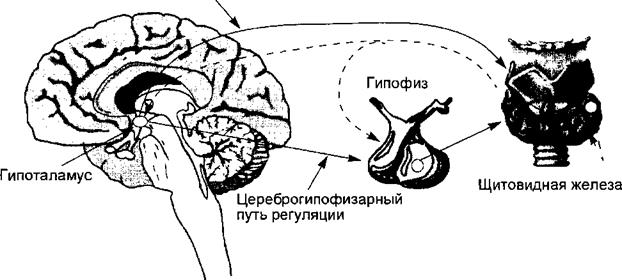
Рис. 3.12. Основные пути регуляции эндокринной системы. Пунктирные стрелки — обратные связи регуляции.
ность тех желез, для которых в гипофизе секретируются специальные тропные гормоны или тропины, например щитовидная железа или кора надпочечников.
Центральной для регуляции эндокринных функций структурой нервной системы является гипоталамус. Этот отдел осуществляет оба пути управления, т. е. и нервный, и гипофизарный. Управляющая функция гипоталамуса связана с наличием здесь групп нейронов, обладающих способностью синтезировать и секретировать специальные регуляторные пептиды — нейрогормоны. Таким образом, гипоталамус является одновременно и нервным, и эндокринным образованием, играя ключевую роль в интеграции нервных и гуморальных механизмов регуляции, осуществляя нейрогумо- ральное управление функциями. Свойство нейронов гипоталамуса синтезировать и секретировать регуляторные пептиды получило название нейросекреции. В принципе, этим свойством обладают все нервные клетки, поскольку нейроны транспортируют синтезированные в них белки, ферменты и другие молекулы с помощью аксонального тока. У нейронов гипоталамуса эта способность приобрела специфические свойства регуляторов эндокринной системы.
Нейросекрет, образующийся в теле гипоталамического нейрона, хранится в нем в виде гранул и путем аксонального транспорта переносится в структуры мозга, ликвор или гипофиз. Соответственно направлению транспорта, в гипоталамусе человека выделены 3 нейроэндокринные системы.
1) Гипоталамо-экстрагипоталамная система представлена нейросекреторными клетками, аксоны которых уходят за переделы гипоталамуса в другие структуры головного мозга — таламус, лимбику, продолговатый мозг — и выделяют нейропептиды, выполняющие медиаторную и модуляторную роль: вазопрессин, эндогенные опиоиды, нейротензин, вещество Р, соматостатин, киоторфин и др.
2) Гипоталамо-аденогипофизарная (переднегипофизарная) система образована пептид- и моноаминергическими нейросекреторными клетками мелкоклеточных ядер заднего гипоталамуса. Аксоны этих клеток образуют контакты в области срединного возвышения на капиллярах (аксовазальные контакты) первичной сети портальной системы гипофиза, в которую и поступают нейропептиды, стимулирующие (либерины) или подавляющие (статины) синтез и секрецию тропных гормонов аденогипофиза. Таким образом, связь гипоталамуса с аденогипофизом осуществляется нейроге- мальным путем (через кровь).
3) Гипоталамо-нейрогипофизарная (заднегипофизарная) система представлена нейросекреторными клетками крупноклеточных ядер переднего гипоталамуса — супраоптического и паравентрикулярного. Аксоны этих клеток опускаются в нейрогипофиз, по ним в заднюю долю гипофиза поступают и депонируются нейропептиды вазопрессин и окситоцин, связанные со специфическими белками нейрофизинами.
Гипоталамические нейропептиды в зависимости от места реализации регуляторного эффекта делят на три группы: 1) висцерорецепторные нейрогормоны, обладающие преимущественным действием на висцеральные органы (вазопрессин, окситоцин), 2) нейрорецепторные нейрогормоны или нейромодуляторы и медиаторы, обладающие выраженными эффектами на функции нервной системы (эндорфины, энкефалины, нейротензин, вазопрессин, ангиотензин и др.), 3) аденогипофизрецепторные нейрогормоны, регулирующие деятельность железистых клеток аденогипофиза (кортиколиберин, соматостатин, тиреолиберин и др.).
Звенья общего и гуморального управления имеют многочисленные обратные связи, контролирующие процессы синтеза и секреции, уровень гормонов в крови и реализацию их эффектов в органах и тканях.
Звено синтеза и секреции гормонов. Особенности синтеза гормонов в эндокринных клетках зависят от химической структуры гормонов. По химической природе все гормоны подразделяют на три группы: 1) производные аминокислот — тиреоидные гормоны, адреналин, гормоны эпифиза; 2) пептидные гормоны, простые (протеины) и сложные (гликопротеиды) белки — гипоталамические нейропептиды, гормоны гипофиза, островкового аппарата поджелудочной железы, околощитовидных желез; 3) стероидные гормоны, образующиеся из холестерина гормоны коры надпочечников, половых желез, гормон почечного происхождения кальцитриол.
Синтез гормонов эндокринными клетками происходит непрерывно, его интенсивность зависит не только от регуляторных сигналов звена управления, но и от величины секреции. Известный в биохимии принцип торможения синтеза конечным продуктом обусловливает подавление образования гормонов при сниженном их удалении из клеток и, напротив, активация секреции повышает синтез гормонов. Таким образом, звенья синтеза и секреции гормонов взаимосвязаны.
Звено депонирования связано с синтезом и секрецией гормонов, поскольку гормоны обычно депонируются в тех же клетках тканей, где образуются. Депонирование гормонов эндокринной тканью может осуществляться в специализированных гранулах (мозговое вещество надпочечников) или в специализированных структурах железы (коллоид фолликулов щитовидной железы). Гормоны депонируются в виде связанных форм с белками, мак- роэргическими фосфатами, нуклеопротеидами или металлами. Однако некоторые гормоны могут депонироваться и в несекреторных тканях, клетками которых они захватываются из крови. Так, например, могут депонироваться катехоламины.
Звено транспорта представлено жидкостями внутренней среды, например кровью, переносящими гормоны как в свободной, так и в связанной формах. Связывание гормонов происходит как с мембранами клеток крови (эритроциты, тромбоциты), так и с белками плазмы крови, при этом активность таких связанных форм крайне низкая, поскольку они плохо проходят через гистогематические барьеры и не могут взаимодействовать со специфическими для них клеточными рецепторами. Свободные (т. е. несвязанные) формы гормонов являются активными, поскольку проходят через барьеры, взаимодействуют с мембранными рецепторами и вызывают физиологические эффекты. Вместе с тем физико-химическая связь гормонов с клетками крови и белками плазмы является формой их депонирования во внутренней среде, поскольку удаление связанных гормонов во внешнюю среду через органы выделения затруднено, а при необходимости гормоны могут освобождаться из связанных форм, переходить в свободную активную форму и вызывать регуляторные эффекты без дополнительной активации их синтеза и секреции.
Транспорт гормонов кровью участвует в реализации механизма обратной связи со звеном управления, поскольку содержащиеся в крови гормоны могут прямо воздействовать на гипоталамус или гипофиз.
Звено метаболизма гормонов требуется не только для разрушения образовавшихся гормонов, но и для уменьшения числа информационных молекул и ослабления их регуляторного эффекта. Метаболические превращения гормонов приводят к образованию новых информационных молекул с отличающимися от основного гормона свойствами, метаболическими и физиологическими эффектами. Метаболизм гормонов осуществляется под влиянием ферментов в самих эндокринных тканях, печени, почках и в тканях-эффекторах. Образование при метаболизме гормонов новых информационных молекул в тканях-эффекторах обеспечивает в них реализацию и новых биохимических и физиологических эффектов. Так, например, дейодирование тироксина в клетках-мишенях ведет к образованию трийодтиронина, обладающего значительно более сильным физиологическим эффектом; метаболизм аланиновой боковой цепи тиреоидных гормонов приводит к образованию тироуксусных кислот с более выраженным эффектом на энергетический обмен клеток. В то же время снижение метаболической деградации молекул гормонов в печени влечет за собой избыточный эффект гормонов на ткани, несмотря на то что эндокринные клетки не повышают секреции гормонов. Многие метаболиты гормонов выделяются из организма с мочой и их концентрации в моче исследуются для оценки функции железы.
Звено выделения гормонов представлено в организме почками, потовыми железами, слюнными железами, желчью и пищеварительными соками. Выведение информационных молекул гормонов и их метаболитов из крови происходит в наибольшей степени через почки с мочой. Выведение гормонов, наряду с депонированием и метаболическим разрушением, защищает организм от избыточности гормональных эффектов.
В звене эффектора гормонов осуществляется реализация биохимических и физиологических эффектов гормональной регуляции. Поступая через жидкую внутреннюю среду к клеткам эффекторов, гормоны связываются со специфическими для них клеточными рецепторами. В связи с этим влияния гормонов не диффузные, предназначенные всем клеткам организма, а строго специфичные, адресованные конкретным клеткам, содержащим рецепторы к определенному гормону. Соответственно, и чувствительность разных тканей к гормональному регулирующему воздействию неодинакова, так как определяется наличием и количеством специфических рецепторов. Ткани, имеющие большое количество рецепторов с высоким сродством к определенному гормону, называют тканями- или органами- мишенями этого гормона. В зависимости от локализации в клетках рецепторы делят на плазматические (расположенные на плазматической мембране клеток), цитозольные (находящиеся в цитоплазме), и ядерные (локализованные в ядре).
3.4.2. Виды и пути действия гормонов
Различают пять видов действия гормонов на ткани-мишени: метаболическое, морфогенетическое, кинетическое, корригирующее и реактогенное.
• Метаболическое действие гормонов вызывает изменение обмена веществ в тканях. Оно происходит за счет трех основных гормональных влияний. Во-первых, гормоны меняют проницаемость мембран клетки и органоидов и транспорт через них субстратов, ферментов, ионов и метаболитов. Во-вторых, гормоны меняют активность ферментов в клетке. В-третьих, гормоны изменяют синтез ферментов, индуцируя или подавляя их образование за счет влияния на генетический аппарат ядра клетки, как прямо вмешиваясь в процессы синтеза нуклеиновых кислот и белка, так и опосредованно через энергетическое и субстратно-ферментное обеспечение этих процессов. Сдвиги метаболизма, вызываемые гормонами, лежат в основе изменения функций клеток, ткани или органа.
Молекула гормона (первичный посредник)
I —
Рецептор клеточной мембраны
Мембранный фермент
Вторичный посредник
Внутриклеточные эффекты

Изменение проницаемости Изменение метаболизма Активация генома
Изменение функции

