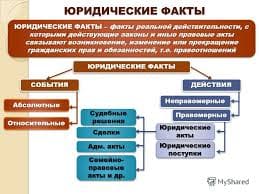
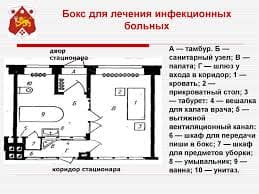
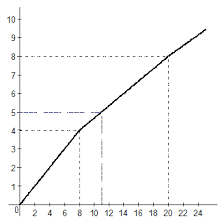
2020-05-25
2020-05-25 186
186
Взаимодействуя с У2-рецептором, вазопрессин обеспечивает как трансцеллюлярный транспорт воды (из внутриканальцевой жидкости через водные каналы аквапоринов-2 апикальной мембраны, микротубулярную систему клеток и через аквапорины-3 и -4 базолатеральной мемеб- раны в интерстициальное перетубулярное пространство), так и парацеллюлярный транспорт воды (через межклеточные плотные соединения)
лу активации, транспорта и встраивания в мембрану, или подвергаются разрушению. Вазопрессин стимулирует всасывание воды и в железах внешней секреции, в желчном пузыре.
Нейропептид вазопрессин поступает по аксонам экстрагипоталамиче- ской системы в другие отделы мозга (лимбика, средний мозг) и участвует в формировании жажды и питьевого поведения, механизмах терморегуляции, в нейрохимических механизмах памяти, формировании биологических ритмов и эмоционального поведения.
Вазопрессин стимулирует секрецию кортикотропина в аденогипофизе, подавляет выделение лютропина при стрессе. Метаболические эффекты вазопрессина заключаются в стимуляции гликогенолиза в печени, стимуляции секреции инсулина, повышении синтеза в печени антигемофиличе- ского глобулина А, продукции фактора Виллебрандта.
Недостаток вазопрессина проявляется резко повышенным выделением мочи низкого удельного веса, что называют «несахарным диабетом», а избыток гормона ведет к задержке воды в организме.
6.2,22 Регуляция секреции и физиологические эффекты окситоцина
Синтез окситоцина в гипоталамических нейронах и его секреция нейрогипофизом в кровь стимулируется рефлекторным путем при раздражении рецепторов растяжения матки и механорецепторов сосков молочных желез. Усиливают секрецию гормона эстрогены. Болевой стресс резко повышает секрецию окситоцина, а этиловый спирт ее угнетает. Возрастает секреция окситоцина и при повышении осмотического давления внеклеточной среды. Несмотря на то что мембранный рецептор окситоцина в клетках-мишенях относится к той же группе, что и У2-рецептор вазопрессина, связывание гормона со специфическим рецептором вызывает снижение в клетках уровня цАМФ. Образование окситоцин-рецепторного комплекса повышает в клетках содержание Са2+ (вторичный посредник) и активирует ионные каналы мембраны, приводя к ее деполяризации.
Основные эффекты окситоцина состоят в стимуляции сокращения матки при родах (чему способствуют высокие концентрации эстрогенов в крови), сокращении гладких мышц протоков молочных желез, что вызывает выделение молока, а также в регуляции водно-солевого обмена и питьевого поведения. Окситоцин является одним из дополнительных факторов регуляции секреции гормонов аденогипофиза, наряду с либеринами. В структурах мозга окситоцин может выступать в роли медиатора или модулятора синаптических процессов, участвовать в механизмах памяти, стимулируя процессы забывания. Окситоцин активирует клеточный иммунитет, оказывает инсулиноподобное действие на жировую ткань. Повышенные количества окситоцина в крови могут вызывать снижение артериального давления.
6.2.5. Гормоны промежуточной доли
Меланотропин у взрослого человека, в отличие от животных с обильным волосяным покровом, практически не синтезируется. Функции этого гормона, заключающиеся в синтезе меланина, его дисперсии в отростках меланоцитов кожи, увеличении свободного пигмента в эпидермисе и, в конечном счете, повышении пигментации кожи и волос, выполняют у человека кортикотропин и липотропин. Эти гормоны, как и меланотропин, образуются из единого предшественника, в связи с чем при избыточной секреции кортикотропина усиливается пигментация кожи. Этому способствуют и близкие гормону пептиды плаценты. Меланотропин тем не менее играет роль как мозговой пептид в нейрохимических процессах памяти.
6.2.4. Эндогенные опиаты
Гипоталамо-гипофизарная система является основным местом синтеза и секреции эндогенных опиатов — полипептидов эндогенного происхождения, обладающих морфиноподобным аналгезирующим действием. Выделяют четыре семейства эндогенных опиатов: эндорфины (а-, /3- и у-), динорфины А и В, неоэндорфины а и Д метионин- и лейцин-энкефалины. В аденогипофизе образуются эндорфины, динорфины и неоэндорфины, тогда как энкефалины синтезируются нейронными структурами разных отделов мозга, прежде всего таламуса и гипоталамуса, и клетками слизистой оболочки кишечника. Опиатные пептиды в центральной нервной системе выполняют роль медиаторов и модуляторов синаптической передачи, обеспечивают функцию противоболевой системы мозга. На мембранах клеток выявлены специфические опиатные рецепторы трех основных типов: ц, 5 и к. Морфин и бета-эндорфин взаимодействуют преимущественно с ц-рецептора- ми, энкефалины — с 5-рецепторами, а динорфины — с к-рецепторами, однако существует и перекрестное взаимодействие. Помимо подавления болевых ощущений эндогенные опиаты участвуют в регуляции адаптивного поведения и реакций организма на стресс, модулируют секрецию гормонов адено- и нейрогипофиза (соматотропина, пролактина, гонадотропинов, вазопрессина). Опиоидные пептиды регулируют процесс всасывания в желудочно-кишечном тракте, угнетают секрецию панкреатического сока и соляной кислоты в желудке. Опиоидные рецепторы выявлены на окончаниях аксонов в мозговом слое надпочечников и симпатических ганглиях, где опиоидные пептиды выполняют роль модуляторов синаптической передачи.