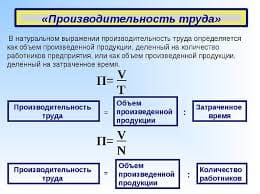
2020-05-25
2020-05-25 202
202Указаны ферменты, контролирующие этапы синтеза молекулы гема в митохондриях, а также хромосомы, ответственные за синтез цепей глобина А (р-цепи) и гемоглобинов плода (у- и 8- цепи). Пояснения в тексте.
Соединение гемоглобина с молекулой О2 называется оксигемоглобином. Сродство гемоглобина к кислороду и диссоциация оксигемоглобина (отсоединения молекул кислорода от оксигемоглобина) зависят от напряжения кислорода (РО2), углекислого газа (РСО2) в крови, pH крови, ее температуры и концентрации 2,3-ДФГ в эритроцитах. Так, сродство повышают увеличение РО2 или снижение РСО2 в крови, нарушение образования 2,3- ДФГ в эритроцитах. Напротив, повышение концентрации 2,3-ДФГ, снижение РО2 крови, сдвиг pH в кислую сторону, повышение РСО2 и температуры крови — уменьшают сродство гемоглобина к кислороду, тем самым облегчая ее отдачу тканям. 2,3-ДФГ связывается с p-цепями гемоглобина, облегчая отсоединение О2 от молекулы гемоглобина. Увеличение концентрации 2,3-ДФГ наблюдается у людей, тренированных к длительной физической работе, адаптированных к длительному пребыванию в горах. Оксигемоглобин, отдавший кислород, называется восстановленным, или дезоксигемоглобином. В состоянии физиологического покоя у человека гемоглобин в артериальной крови на 97 % насыщен кислородом, в венозной — на 70 %. Чем выраженней потребление кислорода тканями, тем ниже насыщение венозной крови кислородом. Например, при интенсивной физической работе потребление кислорода мышечной тканью увеличивается в несколько десятков раз и насыщение кислородом оттекающей от мышц венозной крови снижается до 15 %. Содержание гемоглобина в отдельном эритроците составляет 27,5—33,2 пикограмма. Снижение этой величины свидетельствует о гипохромном (т. е. пониженном), увеличение — о гипер- хромном (т. е. повышенном) содержании гемоглобина в эритроцитах. Этот показатель имеет диагностическое значение. Например, гиперхромия эритроцитов характерна для В12-дефицитной анемии, гипохромия — для железодефицитной анемии.
7.1.3. Старение и разрушение эритроцитов в организме
Эритроциты у человека функционируют в крови максимум 120 дней, в среднем 60—90 дней. Старение эритроцитов связано с уменьшением образования в эритроците количества АТФ в ходе метаболизма глюкозы в этой клетке крови. Уменьшенное образование АТФ, ее дефицит нарушает в эритроците процессы, обеспечиваемые ее энергией, — восстановление формы эритроцитов, транспорт катионов через его мембрану и защиту компонентов эритроцитов от окисления, их мембрана теряет сиаловые кислоты. Старение эритроцитов вызывает изменения мембраны эритроцитов: из дискоцитов они превращаются в эхиноциты, т. е. эритроциты, на поверхности мембраны которых образуются многочисленные выступы, выросты (рис. 7.3).
Причиной формирования эхиноцитов помимо уменьшения воспроизводства молекул АТФ в эритроците при старении клетки является усиленное образование лизолецитина в плазме крови, повышенное содержание в ней жирных кислот. Под влиянием перечисленных факторов изменяется соотношение поверхности внешнего и внутреннего слоев мембраны эритроцита за счет увеличения поверхности внешнего слоя, что и приводит к появлению выростов на мембране.
По степени выраженности изменений мембраны и формы эритроцитов различают эхиноциты I, II, III классов и сфероэхиноциты I и II классов. При старении эритроцит последовательно проходит этапы превращения в эхиноцит III класса, теряет способность изменять и восстанавливать дисковидную форму, превращается в сфероэхиноцит и разрушается. Устранение дефицита глюкозы в эритроците легко возвращает эхиноциты I—II классов к форме дискоцита. Эхиноциты начинают появляться, например, в консервированной крови, сохраняемой в течение нескольких недель при 4 °C, или в течение 24 ч, но при температуре 37 °C. Это связано с уменьшением образования АТФ внутри клетки, с появлением в плазме крови лизолецитина, образующегося под влиянием лецитин-холестерол-ацетилтранс- ферразы, ускоряющих старение клетки. Отмывание эхиноцитов в свежей плазме от содержащегося в ней лизолецитина или активация в них гликолиза, восстанавливающей уровень АТФ в клетке, уже через несколько минут возвращает им форму дискоцитов.
Стареющие эритроциты становятся менее эластичными, вследствие чего разрушаются внутри сосудов (внутрисосудистый гемолиз) или же становятся добычей захватывающих и разрушающих их макрофагов в селезенке, купферовских клетках печени и в костном мозге (внесосудистый или внутриклеточный гемолиз). Внутриклеточным гемолизом в сутки разрушается 80—90 % старых эритроцитов, содержащих 6—7 г гемоглобина, из которых освобождается в макрофагах до 30 мг железа. После отщепления от гемо-

Рис. 7.3. Схема формирования эхиноцитов и стоматоцитов из дискоцита (нормоцита) при разных значениях pH, создаваемых in vitro.
I — сферостоматоцит, II — стоматоцит, III — дискоцит, IV — эхиноцит, V — сфероэхиноцит.
глобина гем превращается в желчный пигмент билирубин, который поступает с желчью в кишечник и под влиянием микрофлоры кишечника последовательно превращается в уробилиноген, а затем в стеркобилиноген. Оба соединения выводятся из организма с калом и мочой, под влиянием света и воздуха превращаясь в стеркобилин и уробилин. При метаболизме 1 г гемоглобина образуется 33 мг билирубина.
Внутрисосудистым гемолизом разрушается 10—20 % эритроцитов. При этом гемоглобин поступает в плазму, образует с плазменным гликопротеином гаптоглобином комплекс гемоглобин—гаптоглобин. В течение 10 мин 50 % комплекса поглощается из плазмы паренхиматозными клетками печени, что предупреждает поступление свободного гемоглобина в почки и тромбирование им их нефронов. У здорового человека в плазме содержится около 1 г/л плазмы гаптоглобина, что оставляет несвязанным с ним в плазме крови не более 3—10 мг гемоглобина. Молекулы гема, высвобождающиеся из связи с глобином при внутрисосудистом гемолизе, связываются белком плазмы — гемопексином, транспортируются им в печень и также поглощаются паренхиматозными клетками печени, где подвергаются ферментному разрушению до билирубина.
7.1.4. Роль ионов железа в эритропоэзе
В организме содержится 4—5 г железа в виде резервного ('/<) и функционально-активного железа (3/4). 62—70 % железа находится в гемоглобине эритроцитов, 5—10 % — в миоглобине, остальное — в тканях, где оно участвует во многих метаболических процессах: в составе металсодержащих энзимов—цитохромов обеспечивают митохондриальный транспорт электронов, синтез ДНК и деление клеток, метаболизм гормонов мозгового вещества надпочечников, детоксикационные механизмы, снижающие активность токсичных веществ под влиянием цитохрома Р450, в состав которого входит железо; поддерживают активность глицерол-3-фосфатдегидрогеназы в поперечно-полосатых мышцах, что сохраняет физическую работоспособность человека. Например, у людей, страдающих дефицитом железа, физическая работоспособность резко снижается, но восстанавливается после приема препаратов железа.
Резервное железо постоянно переходит в функциональное и обратно. Так, за счет повторного использования костный мозг получает ежесуточно 20—25 мг железа. Ионы железа (Fe++) ежедневно всасываются в кишечнике из пищи. Fe2+ в кишечном содержимом вначале связывается с белком- рецептором на поверхности эпителия слизистой оболочки кишки (р3-ин- тегрином), что необходимо для перехода его через мембрану клетки слизистой в цитозоль. Далее, образовав комплекс с цитозольным транспортным белком- мобилферрином, Fe2+ пересекает клетку эпителия слизистой кишечника оставаясь в этом комплексе вплоть до поступления Fe2+ в капиллярную сеть кишечника. С выходом в кровоток двухвалентное железо окисляется до трехвалентного белком плазмы крови церрулоплазмином и присоединяется к трансферрину, гликопротеину плазмы крови. Трансферрином Fe3+ доставляется к тканям и используется в митохондриях эритробластов для синтеза гема, депонируется в макрофагах в виде резерва. Плазма содержит от 1,8 до 2,6 мг/л трансферрина, 1 мг которого связывает 1,25 мкг Fe. В общем объеме плазмы содержится около 3 мг Fe2+. В норме лишь 7з трансферрина плазмы насыщена железом. Количество железа, которое может быть связано трансферрином, называется общей железосвязывающей способностью крови и в норме составляет 250—400 мкг % (45— 72 мкмоль/л). Концентрация железа в сыворотке крови у взрослых людей обоего пола колеблется от 50 до 160 мкг % (9—29 мкмоль/л). С мочой выводится за сутки 60—100 мкг железа.
Комплекс трансферрин—железо фиксируется на рецепторах мембран эритробластов. Количество этих рецепторов уменьшается за счет торможения их синтеза в ходе созревания эритроидных клеток и исчезает после созревания ретикулоцитов, поэтому в зрелые эритроциты железо не включается. Освобождение железа из комплекса трансферрин — железо обеспечивается энергией молекул АТФ, образуемых в эритробластах. Молекула трансферрина, отдавшая железо, смещается с мембранного участка молекулами трансферрина, связанными с железом, поскольку их сродство к рецепторам более сильное. Железо, поступившее в эритробласт, используется в митохондриях для синтеза гема и депонируется в эритробласте в виде резерва. В макрофагах печени и костного мозга резервное железо депонируется в молекуле ферритина. Внутри лизосом молекулы ферритина образуют большие аморфные нерастворимые агрегаты — гемосидерин. Таким образом, ферритин и гемосидерин — это формы резервного железа в клетках. Из клеточного резерва железо освобождается в двухвалентном состоянии (благодаря энзиму ксантиноксидазе, аскорбиновой кислоте и др.), затем церулоплазмин окисляет Fe2+ до трехвалентного состояния, Fe3+ соединяется с трансферрином и транспортируется с плазмой крови к эритробластам.
Абсорбции железа эпителиальными клетками желудочно-кишечного тракта способствуют сниженное насыщение трансферрина железом и повышенная эритропоэтическая активность крови. Абсорбция снижается при увеличении концентрации железа в клетках слизистой оболочки кишечника. В кишечнике более эффективна абсорбция Fe2+, чем Fe3+. Поэтому аскорбиновая кислота, фруктоза, аминокислоты (цистеин, метионин) поддерживают двухвалентную форму железа и ускоряют его абсорбцию. В кишечнике лучше абсорбируется биодоступное железо, входящее в состав гема (мясные продукты, кровяная колбаса), чем железо из пищи растительного происхождения. Абсорбция Fe2+ в желудочно-кишечном тракте зависит от возраста человека, функционального сотояния его организма. Она наиболее высокая у детей первых месяцев жизни и достигает 57 %, у 7— 10-летних — 7,75—17,75 %, у взрослых мужчин и женщин — 1,1—11,2 % и у беременных женщин — 20 % от поступившего в желудочно-кишечный тракт железа. Отсюда ежедневная потребность в железе, абсорбируемом из пищи в организме человека, составляет: у детей от 0 до 4 мес жизни — 0,5 мг/день, от 1 года до 12 лет — 1,0 мг/день, женщины детородного периода— 2,8 мг/день (во время менструации потери железа составляют от 5 до 45 мг), молодые мужчины — 1 мг/день, старики — 0,9 мг/день, беременные женщины — 3—3,5 мг/день. В организм последних за время беременности должно поступать до 1 г железа (500 мг железа используется для синтеза дополнительного количества гемоглобина в организме беременной, 300 мг Fe2+ необходимо формирующемуся плоду и 200 мг Fe2+ компенсируют естественные потери металла организмом беременной женщины). Дефицит железа в организме человека приводит к развитию гипохромной анемии.
7.7.5. Эритропоэз
Эритропоэз — процесс образования эритроцитов в костном мозге. Первой клеткой эритроидного ряда, образующейся из колониеобразующей клетки эритроцитарной (КОК-Э) — клетки-предшественницы эритроидного ряда, является проэритробласт, из которого в ходе 4—5 последующих удвоений и созревания образуется 16—32 зрелых эритроцита. Схема процесса: 1 проэритробласт (удвоение) два базофильных эритробласта I порядка 4 базофильных эритробласта II порядка 8 полихроматофильных эритробластов I порядка 16 полихроматофильных эритробластов II порядка -> 32 полихроматофильных нормобласта 32 оксифильных нормобла-