 2020-05-25
2020-05-25 150
150
Под влиянием паратирина почка се <ретирует кальцитриол, основные эффекты которого (жирные стрелки) заключаются в стим} пяции всасывания в кишечнике в кровь ионов кальция и фосфата и усилении их захвата кост ной тканью. Эффекты кальцитриола на почку (стимуляция реабсорбции кальция и фосфора) в сражены слабее.
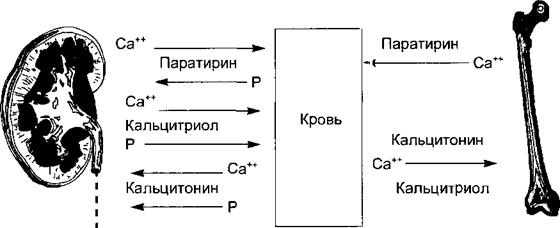

Рис. 6.26. Эффекты трех кальцийрегулирующих горм >нов на органы-мишени.
В почке паратирин и кальцитриол активируют реабсорбции кальция, а кальцитонин ее угнетает. И паратирин, и кальцитонин подавляют реабсорбцию < юсфата. В кишечнике кальцитриол и паратирин активируют всасывание кальция и фосфата Кальцитонин и кальцитриол способствуют отложению кальция в костях, а паратирин актп шрует резорбцию кости и выход кальция в кровь.
лирование происходит по 24-му атому углерода и синтезируется второе соединение — 24,25-(OH)2-D, которое обладает си >собностью угнетать секрецию паратирина по принципу обратной связи. Инактивация кальцитриола происходит в печени.
Основной эффект кальцитриола (рис. 6.25) заключается в активации всасывания кальция в кишечнике. Гормон стих улирует все три этапа всасывания: захват ворсинчатой поверхностью клетки, внутриклеточный транспорт, выброс кальция через базолатеральн 'Ю мембрану во внеклеточную среду. Действие кальцитриола на эпителиальные клетки кишечника состоит в индуцировании синтеза энтероцитаии специальных кальций- связывающих и транспортирующих белков — к алъбайндинов, Кальцитриол повышает в кишечнике и всасывание фосфато и Почечные эффекты гормона заключаются в стимуляции реабсорбции оосфата и кальция канальцевым эпителием. Эффекты кальцитриола на костную ткань связаны с прямой стимуляцией остеобластов и обеспечением костной ткани усиленно всасывающимся в кишечнике кальцием, что активирует рост и минерализацию кости. Эффекты кальцитриола, как и зсех стероидных гормонов, делятся на геномные и негеномные. Геномные эффекты обусловливают синтез кальбайндинов, активирование остеобла< тов и синтеза костной ткани. Увеличение транспорта кальция внутрь клегок, например в скелетных и сердечной мышцах, в остеобластах, энтероцгтах, гепатоцитах и клетках
околощитовидных желез, происходит быстро и обусловлено негеномным действием гормона. Под влиянием гормонрецепторного мембранного комплекса в клетках происх< щит образование вторичного посредника диацил- глицерола и активация протеинкиназы С. Кальцитриол меняет в клетке и уровень цАМФ и цГМФ, что ведет к модификации геномного эффекта. Наличие специфических рецепторов к гормону у многих клеток тканей (в молочной железе, эндокринных железах, нервной системе), способность кальцитриола активиров; ть транспорт кальция в большинстве из них свидетельствуют о широком спектре эффектов этого гормона.
Участие трех кальцийрегулирующих гормонов в гомеостазисе кальция и фосфора показано на рис. 6.26.
Недостаточность кальцитриола проявляется в виде рахита, т. е. нарушения созревания и кальцификации хрящей и кости у детей, либо остеомаляции, т. е. падения минер ишзации костей после завершения роста скелета. При этом сдвиги уровня кальция в крови и клетках обусловливают угнетение нейромышечной возбудимости и мышечную слабость.
6.8.3.2. Образование ренина и основные функции ренин-ангиотензи н-алъдостероновой системы
Ренин образуется в виде г роренина и секретируется в юкстагломерулярном аппарате (ЮГА) (от латинских слов juxta — около, glomerulus — клубочек) почек миоэпителиоидны ии клетками приносящей артериолы клубочка, получившими название юкстагломерулярных (ЮГК). Структура ЮГА приведена на рис. 6.27. В ЮГА кроме ЮГК также входит прилегающая к приносящим артериолам час ь дистального канальца нефрона, многослойный эпителий которого образ} ет здесь плотное пятно — macula densa. Секреция ренина в ЮГК регулируется четырьмя основными влияниями. Во-первых, величиной давления крови в приносящей артериоле, т. е. степенью ее растяжения. Снижение растяжения активирует, а увеличение — подавляет секрецию ренина. Во-втоэых, регуляция секреции ренина зависит от концентрации натрия в моче дистального канальца, которая воспринимается macula densa — своеобразным Na-рецептором. Чем больше натрия оказывается в моче дистального канальца, тем выше уровень секреции ренина. В-третьих, секреция ренина регулируется симпатическими нервами, ветви которых заканчиваются на ЮГК, медиатор норадреналин через бета-адренорецепторы стимулирует секрецию ренина. В-четвертых, регуляция секреции ренина осуществляется по механизму отрицательной обратной связи, включаемой уровнем I крови других компонентов системы — ангиотензина и альдостерона, а т юсе их эффектами — содержанием в крови натрия, калия, артериальным давлением, концентрацией простагландинов в почке, образующихся под влиянием ангиотензина.
Кроме почек образование ренина происходит в эндотелии кровеносных сосудов многих тканей, м гокарде, головном мозге, слюнных железах, клубочковой зоне коры надпочечников.
Секретированный в кр >вь ренин вызывает расщепление альфа-глобули- на плазмы крови — ангиотензиногена, образующегося в печени. При этом в крови образуется (рис. 6.'.’.8) малоактивный декапептид ангиотензин-1, который в сосудах почек, легких и других тканей подвергается действию превращающего фермента (карбоксикатепсин, кининаза-2), отщепляющего от ангиотензина-1 две аминокислоты. Образующийся октапептид ангиотензин-11 обладает большим числом различных физиологических эффектов, в том числе стимуляцией К1убочковой зоны коры надпочечников, секрети-
 |

импатическая активация
I
—► ЮГК —Macula —NaCI
—— densa
Ренин
Ангиотензиноген 1 2 3 4 5 6 7 8 < 10 V11 12 13 14
Asp-Arg-Val-Tyr-lle-His-Pro-Phe-Hs-Leu-Leu-Val-Tyr-Ser-R
Ангиотензин-I 1 23 4567 8 9 10
Asp-Arg-VakTyr-lle-His-Pro-Phethbs-Leu
I— Дипептидкарбоксилаза (ПФ)
Ангиотензин-Ц 1 234567 8
Asp-Arg-Val-Tyr-lle-His-Pro-Phe








