 2014-02-03
2014-02-03 7107
7107Крупный рогатый скот. В кариотипе его содержится 60 хромосом. Впервые их подсчитал Краллингер в 1927 г. Обнаружены разные формы числовых и структурных аномалий кариотипа, которые сочетаются с нарушением плодовитости, эмбриональной смертностью, интерсексуальностью, злокачественными процессами (лейкоз, саркома и др.), врожденными уродствами и некоторыми генетическими аномалиями обмена веществ, болезнями
животных.
Числовые аномалии кариотипа (анеуплоидия). Числовые аномалии хромосом относят к вновь возникающим мутациям. Однако имеются исследования, которые показывают, что может быть семейная предрасположенность к анеуплоидии. Так, Герцог, Хен и Олишлегер при описании шести случаев трисомии по 17-й хромосоме (новой форме трисомии у крупного рогатого скота), сочетающейся с синдромом общего недоразвития телят (нанизм), гидроцефалией, микрофтальмией, аномалиями сердца и крипторхизмом, указывают на генетическую предрасположенность к нерасхождению хромосом.; С. Г. Куликова (1991) обнаружила трисомию по 19-й паре хромосом, которая ассоциировалась с прогнатией нижней челюсти у теленка (рис. 58).
Гаметы с трисомией, моносомией, нуллисомией и полисо-мией обычно вызывают летальный исход уже на ранних стадиях эмбрионального развития и являются продуктом нарушения спермио- или овогенеза у носителей транслокаций. После рождения наблюдают числовые нарушения только по мелким ауто-сомам и половым хромосомам.
Полиплоидия. Увеличение числа наборов хромосом — полиплоидию наблюдали как в соматических, так и в половых клетках в мозаичной форме, т. е. как определенный процент клеток с аномальным набором хромосом.


Р«. 58. Трнсомия щ• W-Й^ромосоме у чуш,»*, р„гатого «^ (KEG-окряска хромосом)
Полностью (100 %) полипловдные эмбрионы у крупного оо-гатого скота и других видов животных, £ исключ^ием ^таи отмирают на ранних стадиях онтогенеза. Поли^вдию обн^у' живали в бластоцитах телок, забитых через 1™Тд£»й по?лё случки. И. Л. Голвдман и др. отмечали возрастание полижи дни у крупного рогатого скота, больного лейкозов ПодХьш болгарских ученых, бычки с высоким уровней? соматической полиплоидии во взрослом состоянии покажи кТини^еск^^кад тину лейкоза. По мнению ученых, подбор жи™^ c^rSL стабильности кариотипа может способствов^ь эфф^кти^и мероприятий, направленных на искоренение лейкоза у крупного рогатого скота.
Привлекает внимание работа Г. К. Исаковой, В. И. Евсикова, Д. К. Беляева (1976) на норках. В ней они показали, что 11,9 % имплантированных эмбрионов имеют триплоидию, мозаицизм 2п/3п или 2п/4п. Анализ полиплоидии в сперматогенном эпителии самцов норок с неудовлетворительной воспроизводительной характеристикой показал статистически достоверное повышение частоты полиплоидных клеток по сравнению с соответствующим показателем у животных с нормальной плодовитостью (5,9 и 9,6 % соответственно).
Анализ полиплоидии в костном мозге самок и самцов с различной воспроизводительной характеристикой также показал статистически достоверные различия по уровню полиплоидии: частота полиплоидных клеток у самок и самцов с нормальной плодовитостью составила 0,5 и 0,7 % соответственно, у животных с неудовлетворительной воспроизводительной характеристикой (давших в приплоде в 2 раза меньше телят) — 0,9 и 1,4 %.
Высокий процент полиплоидных клеток наблюдали у крупного рогатого скота с раздвоенным крупом — доппельленднеров с врожденным пояснично-крестцовым уродством телят, где частота полиплодии была выше 10 % (максимально 40 %) (А. И. Жи-гачев, 1979; Герцог, Хен, Файнас, 1983).

Pec. 59. Трянслокашш мехцу 1-й н 29-й аутосомамн
Структурные мутации хромосом. Транслокации. Наибольшее количество исследований у крупного рогатого скота проведено по изучению частоты и влияния на плодовитость центрического слияния — транслокации м]ежду 1-й и 29-й аутосомами (рис. 59). Эта аберрация обнаружера'в молочных, мясных и комбинированных породах во многих странах мира, в том числе у голштино-фризской (США, Англия), немецкой черно-пестрой (Германия), айрширской (Швеция), симментальской (Швейцария, Германия, Австрия, Венгрия, Россия, Югославия, Новая Зеландия и др.), монбельярдской (Франция), швицкой (США и другие страны), шароле (Франция), лимузин (Франция, Англия). Еще в 1977 г. насчитывали 28 пород, в которых была обнаружена транслокация 1/29 хромосом, а к 1991 г. уже было 50 пород.
Частота транслокаций при обследовании пород была неодинаковой и составила у швицкой, айрширской пород — 12,8 %, у симментальской в Англии — 4,8, в бывшем СССР — 5 % у быков на племпредприятиях, 10 — у коров и 18 — у ремонтных быков (А. И. Жигачев и др.), у шароле в Англии — 12,8, у лимузинов во Франции — от 4 до 14 %.
Транслокация 1/29 хромосом снижает плодовитость крупного рогатого скота, по отдельным расчетам, на 3,5—10 % и выше. Причины снижения плодовитости связаны с тем, что у гетерозиготных носителей робертсоновской транслокации образуются гаметы о несбалансированным набором хромосом. Так, при носи-тельстве транслокации 1/29 хромосом возможно образование шести типов гамет. Из них 1-й и 2-й типы — это гаметы с избытком, а 4-й и 5-й — с недостатком генетического материала. Использование производителя с кариотипом 2n=59, XY Т 1/29 на коровах с нормальным набором хромосом 2п=60, XX может привести к формированию нежизнеспособных эмбрионов с три-сомией и моносомией по 1-й и 29-й аутосомам. Такие же результаты возможны и при других вариантах скрещиваний.
Коровы — носители транслокации 1/29 хромосом, по данным Густавссона, имеют более низкую молочную продуктивность, поэтому их раньше выбраковывают.
Во многих странах в законодательном порядке запрещено использовать быков — носителей транслокации 1/29 хромосом на станциях искусственного осеменения. Приняты ограничения или требования о цитогенетической аттестации при импорте и экспорте животных или их гамет.
Кроме транслокации у крупного рогатого скота описаны центрические слияния между другими парами аутосом (табл. 48).
48. Типы центрических слияний (транслокаций)
между различными парами аутосом у крупного рогатого скота
(по Густевссону, с нашими дополнениями)
| Пара аутосом, вступившие в центрические слияния | Порода | Страна |
| 1/29 Мозаика 13/21/норма 11/12/15/16, или 13/21, или 14/20 7/11/20/25 5/6/15/16 или 6/16 | Разные Голштино-фризская Симментальская Белая аквитанская и лимузин Декстерская | Разные Венгрия Новая Зеландия, Англия, Венгрия Франция США |
Продолжение
| Пара аутосом, вступившие в центрические слияния | Порода | Страна |
| 2/4 | Британская фризская | Англия |
| 27/29 | Гернзейская | Канада |
| 1/25 | Симментальская | Германия |
| 3/4 | Лимузин | Франция |
| 8/9 | Швицкая | Швейцария |
| 25/27 | Альпийский скот | » |
Сведений о влиянии этих типов слияний на фенотип накоплено недостаточно, за исключением транслокации 25/27, которая снизила плодовитость животных. Рассмотрим на двух конкретных примерах воздействие транслокаций 1/29 и 25/27 хромосом на воспроизводительную функцию коров. Так, итальянские ученые сравнивали показатели воспроизводительной функции и продуктивность коров — полусестер по отцу — носительниц транслокаций и нормальных особей серой альпийской породы по средним показателям. Швейцарские ученые такой же анализ провели на симментальской породе, при этом получили следующие результаты (табл. 49).
49. Число осеменений на зачатие (по Ценеру и др.)
| Порода и кариотип | Число осеменений | Число стельных животных | Индекс осеменений | Разница |
| Серая альпийская | ||||
| 2п=60, XX | 1,18 | 0 16 | ||
| 2п=59, XX, Т 25/27 | 1,34 | |||
| Симментальская (шифр быка) | ||||
| Н2п=60, XX | 1,51 | 0 IS | ||
| Н2п=59, XX, Т 1/29 | 1,66 | U,lJ | ||
| 2п=60, XX | 1,51 | 0,12 | ||
| 2п=59, XX, Т 1/29 | 1,63 |
Для зачатия у коров — носительниц транслокации 25/27 хромосом требовалось большее количество осеменений, чем у их нормальных полусестер. Число дней от отела до последующего плодотворного осеменения (сервис-период) у коров — носительниц транслокаций было выше, чем у их нормальных полусестер (табл. 50).
50. Продолжительность сервис-периода
| Порода и кариотип | Число животных | Сервис-период (дней) | Разница (дней) |
| Серая альпийская | |||
| 2п=60, XX | 75,5 | 9,9 | |
| 2п=59, XX, Т 25/27 | 85,4 | ||
| Симментальская (шифр быка) | |||
| Н2п=60, XX | 74,7 | 23,7 | |
| Н2п=59, XX, Т 1/29 | 98,4 | ||
| 2п=60, XX | 106,0 | 6,5 | |
| 2п=59, XX, Т 1/29 | 112,5 |
Венгерский ученый Ковач (1982) указывает на то, что различия по степени влияния разных типов центрических слияний на воспроизводительную функцию могут обусловливаться неодинаковым уровнем смерти несбалансированных гаплоидных клеток или эмбрионов. Эти различия также могут быть связаны с утратой центромерных участков хромосом, вступающих в транслокацию, или потерей их функциональной активности.
Кроме транслокаций по типу центрических слияний у круп
ного рогатого скота обнаружены также реципрокные транслока
ции и тандемного типа. Так, Хансен (1970) зарегистрировал тан-
демную транслокацию 1-й и 9-й хромосом у датского молочного
скота. Эта аберрация была связана с повышенной эмбриональ
ной смертностью и снижением плодовитости животных пример
но на 10 %. '
Герцог (1972) наблюдал тандемную транслокацгао 1-й и 7-й хромосом у животных немецкой красной породы с гипоплазией левой «f асти большого полушария мозга, расщеплением позвоночника и сегментной аплазией спинного мозга.
Инверсии. Перицентрическая инверсия в 14-й паре хромосом обнаружена Попеску у нормандской породы, шароле и гернзеев. Аберрация заметно снижала плодовитость животных.
Интересные работы по изучению причин нарушения спермиоге-неза и плодовитости быков провел Кнудсен. При анализе гермента-тивного эпителия быков с уменьшенной плодовитостью ученый обнаружил у трех животных транслокации, а у восьми — инверсии. На микрофотографиях, изготовленных при помощи электронного микроскопа, в стадии пахитены была видна петля инверсии, а в стадии поздней анафазы мейоза был виден инверсионный мост, образовавшийся из децентрической хромосомы. Ненормальное поведение хромосом первичных сперматоцитов во время мейоза вследствие инверсии было причиной бесплодия быков.
Делеции, нехватки, поломки хромосом. Утраты средних участков хромосом (делеции) и концевых участков (нехватки) вызывают обычно летальный эффект на ранних стадиях онтогенеза. Их находят также у животных с различной патологией.
Нередко в кариотипе обнаруживают поломки хромосом — хроматидные и хромосомные разрывы с образованием фрагментов генетического материала. Из множества работ по данному вопросу следует выделить исследования Хелнан (1982), который показал, что мелкие делеции или вторичные перетяжки хромосом и изохроматидные разрывы, как он затем их назвал, наследуются и имеют связь с хромотой у крупного рогатого скота вследствие тазобедренных артритов.
Высокая частота вторичных перетяжек обнаружена и в наших исследованиях (А. И. Жигачев и др., 1983) у отдельных животных с врожденными аномалиями и у некоторых быков зарубежного происхождения. Герцог, Хен и Рикк (1977) при обследовании телят черно-пестрой немецкой породы, больных паракерато-зом, установили, что у их отцов и матерей число хромосомных разрывов аутосом было достоверно выше (11,1 и 9,5 %), чем в среднем у взрослых животных (1,4 %). Авторы предлагают использовать число хромосомных разрывов как маркер гетерози-готности по наследственному паракератозу.
Высокую частоту хромосомных разрывов обнаружили у животных, пораженных лейкозом. Делеции, затрагивающие половую Х-хромосому, наблюдали в кариотипе коров с низкой опло-дотворяемостью.
В наших исследованиях у коров с многократными перегулами также отмечены повышенная частота разрывов хромосом и другие аберрации по сравнению с их сверстницами, которые оплодотворялись после первого осеменения.
Из цитированных работ видно, что структурные изменения хромосом — это дополнительная информация о роли генотипа в патологии животных. Вместе с тем возникновение разрывов хромосом может быть индуцировано вирусами и другими тератогенными факторами, что необходимо учитывать при цитогенетичес-ком анализе. Так, делеции, нехватки и пробелы хромосом с высокой частотой отмечены Т. В. Богачевой при анализе влияния на генетический аппарат быков супердоз витаминов А и D.
Хромосомные аномалии могут широко распространиться в породе через производителей, используемых в воспроизводстве, особенно если их спермой осеменяют коров племенных заводов, которые продают ремонтных быков на племпредприятия по искусственному осеменению. Из этого следует вывод о необходимости цитогенетического контроля за распространением хромосомных аномалий в скотоводстве и браковки животных с нарушением кариотипа.
Свиньи. Нормальный кариотип свиньи состоит из 38 хромосом. Впервые его описал Краллингер (1931).
У свиней наблюдаются различные формы аберраций. Наиболее часто у них обнаруживают реципрокные транслокации между различными парами аутосом (рис. 60). Анализ показал, что реципрокные транслокации снижают плодовитость свиней (табл. 51), а также продуктивные качества (среднесуточный прирост массы, признаки мясности и др., табл. 52).
Общее число реципрокных транслокаций у свиней более 20, они снижают жизнеспособность потомков от 25 до 50 %.
Причина уменьшения плодовитости у носителей транслокаций — нарушение мейоза. В процессе мейоза у животных — носителей транслокации образуются гаметы с несбалансированным набором хромосом, которые участвуют в оплодотворении. Так, у четырех гетерозиготных хрячков и свинок — потомков хряка
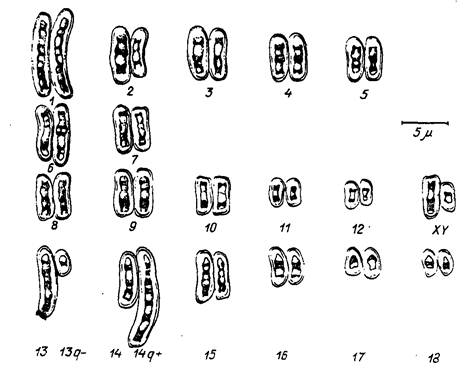
Рис. 60. Транслокация между 13-й н 14-й хромосомами свиньи (но Хагельдорну и др.)
51. Частота вовлачания хромосом свинай в спонтанные раципрокные
транслокации
| Хромосома | Число транслокаций | Транслокации |
| 1-Я | гср (1р • 6q+), rcpjlp-; I4q+), гср+(1р-; 16р+), rep (lq; 15q), rep (Iq; 17q+), rep (lq+; 14q-), rep (lq+; | |
| 2-я | 7q), rep (lp-; 8ql | |
| 3-я 4-я | rep (3p+; 7q~) rep (4q; 14p), rep (4q+; 13q), rep (4q; 15p+) | |
| 5-я | + rcp (5q; 8q+), rep (5p~; 14p+) | |
| 6-я 7-я 8-я | 3 4 | rep (6p; 15a), rep (6p; 14q~), rep (lp-; 6q+) rep (7q; llq), гср (7р*; 13q~), гср (Зр^; 7~), rep ■ Oq; 7q). rep (lp. 8q+) |
| 9-я 10-я | 1 0 | rep (9p+; llq") |
| . 11-я 12-я | 3 0 | rep (llp+; 15q~), rep (7q~; llq+), rep (9p+; llq-) ^ |
| 13-я | rep (llp+; 15q-), rep Qq"; llq+), rep (9p+; llq-) | |
| 14-я | rep (13q^; 14q), rep (lp+; I4q-), rep (4p+; 14q-), rep (lq; 14q), rep (4q~; 14p+), rep (5p~; 14p) | |
| 15-я 16-я 1 Т | rep (llp+; 15q~), rep (15q+; 16q"), rep (16q+; I7q") rep (lp; {6p), rep (lSq*; I6q-), rep (^q^; 17q~) | |
| 17-Я | rep (lq; 17q+), rep (16q+; 17q~) |
52. Срввнение снижения воспроизводительной способности
хряков-носителей реципрокных транслокаций и эмбриональной
смертности у их потомстве
| Транслокация | Снижение размера помета, % | Частота несбалансированных кариотипов, % | Эмбриональная смертность, % |
| гср (lip"1"; 15q-) rcp(13q-; 14p+) rep (4q"; 14p+) ГСР (9p+; llq-) | 34 42 49 50 | 10,6 30,5 38,8 29,9 | 37,2 68,8 65,5 67,7 |
шведской йоркширской породы, гетерозиготного по транслокации (13q~; 14q+), на стадии диакинез — метафаза I наблюдали 17 бивалентов, по одному квадриваленту. У свинок наблюдали хромосомы с кольцевой конфигурацией, что характеризует реци-прокную транслокацию. У хрячков кольцевую конфигурацию наблюдали примерно в 20 % клеток на стадии диакинез — метафаза I. Образование квадривалентов приводит к формированию несбалансированных гамет на стадии метафазы II, что является причиной нарушений процессов эмбриогенеза после оплодотворения. Получены четкие доказательства участия гамет с несбалансированным кариотипом в оплодотворении, что свидетельствует о неэффективности или отсутствии презиготического отбора (элиминации гамет с несбалансированным кариотипом до оплодотворения).
Исследовали предимплантационные эмбрионы, полученные после случки гетерозиготных по реципрокной (13q~; I4q+) транслокации хряков с 10 нормальными свинками. В 36 карио-типированных эмбрионах 14 имели нормальный сбалансированный набор хромосом, 11 — сбалансированный, но гетерозиготный по транслокации, 11 — несбалансированный. В последнем случае у эмбрионов кроме структурной перестройки — транслокации были выражены и изменения числа хромосом в кариоти-пе — трисомия или моносомия как следствие нарушения их расхождения в мейозе.
Во втором варианте исследования случали хряка с нормальным набором хромосом с 10 гетерозиготными свинками. Цитогенетический анализ проведен у 33 эмбрионов. Из них 10 оказались с нормально сбалансированным кариотипом, 14 гетерозиготных сбалансированных, 9 имели несбалансированный набор хромосом. В третьем варианте хряка случали с тремя нормальными свинками. В результате анализа выявлено 17 нормальных сбалансированных и 12 сбалансированных, но гетерозиготных эмбрионов. В четвертом варианте после случки нормального хряка и четырех гетерозиготных свинок изучено 17 эмбрионов, из которых 5 имели нормальный кариотип, 10 оказались гетерозиготными сбалансированными и 2 дегенерированными. В последнем варианте гетерозиготного хряка случали с гетерозиготной свинкой. Из четырех проанализированных эмбрионов 2 имели сбалансированный с гетерозиготной формой транслокации кариотип и 2 эмбриона были дегенерированные.
Эмбриональная смертность, вычисленная по числу желтых тел и живых имплантированных эмбрионов, у гетерозиготных свинок, слученных с нормальными хряками, а также у нормальных и гетерозиготных свинок, слученных с гетерозиготными хряками, составила соответственно 72,3; 68,0; 85,5 %. Таким образом, у хряков и свинок — носителей транслокации хромосом наблюдаются нарушения процессов мейоза и гаметогенеза. Такие спер-мии и яйцеклетки, участвуя в оплодотворении, дают нежизнеспособных эмбрионов, что выражается в резком снижении количества поросят в пометах.
Рещргрокную транслокацию Т (llp+; 15q~) в гетерозиготном состоянии обнаружил Кинг у двух хряков — отца и сына — со сниженной на 56 и 34 % оплодотворяющей способностью. Эта транслокация в сбалансированной форме обнаружена у 41 % живых потомков сына. Несбалансированный кариотип выявлен у 11 % эмбрионов, изученных в период от 10-го до 88-го дня беременности. Эти эмбрионы, несомненно, погибают. 100%-ную эмбриональную смертность наблюдали Бойтерс и др. в потомстве при использовании хряка — мозаика по реципрокной транслокации между 6-й и 15-й хромосомами. Кариотип этого животного был 38, XY/38, XY+ (6q+; 15q~). Высокую степень смертности, включая мертворожденных поросят и потомков с врожденными уродствами, зарегистрировали Попеску и Легатт у хряка, в кариотипе которого обнаружено 24 % клеток со структурными перестройками.
Подтверждением того положения, что снижение плодовитости у свиней может быть связано с нарушением генома, является сообщение Фогта и др. Два хряка-брата, дающих потомство с 50%-ным уменьшенным размером помета, в сравнении с другими хряками и теми же свиноматками оказались миксоплоида-ми - 37, XY- 18/38, XY/39, XY, +18 и 37, XY, - 18/38, XY. При анализе кариотипа их потомков у двух свиней-дочерей с пониженной плодовитостью был обнаружен аномальный набор хромосом.
Норби с соавт. при анализе бластоцист у свиней в 10' % случаев из 38 обнаружили количественные и структурные аномалии, в их числе 4 триплоида, 3 тетраплоида, 1 диплоид/триплоид и одна делеция. При изучении 13 хряков с пониженной плодовитостью и 100%-ной эмбриональной смертностью потомства Бойтерс установил, что основная причина нарушения эмбриогенеза — аномалии кариотипа: транслокации, палочковидные хромосомы, которые наблюдались в 25 % из 40 % эмбрионов, созревших до 25 дней.
В исследованиях М. Л. Кочневой и Н. А. Осиповой (1995) показано увеличение соматической хромосомной нестабильности у поросят с врожденными аномалиями и у свиноматок с низкой
плодовитостью.
Цитогенетический анализ у свиней дал возможность изучить связь нарушений плодовитости с аномалиями в системе половых хромосом. Так, на метафазных препаратах поросят-интерсексов был обнаружен химеризм XX/XY и XX/XXY. Такая же аномалия хромосом в лимфоцитах найдена у плодовитой свиноматки (среднее число поросят в один помет — 14), у которой, по данным Кристенсена, 24 % клеток имели половые хромосомы XY, остальные содержали ХХ-хромосомы. У свиней обнаружена аномалия кариотипа, сходная с синдромом Клайнфельтера у человека и ранее описанная на других видах (2n-39,XXY). Для характеристики спермиогенеза у этого животного исследовали суспензию эпидидимальных клеток и семенники. В содержимом эпидидимуса спермин обнаружены не были. Гистологический анализ, проведенный Ханкоком, показал отсутствие спермато-генного эпителия.
У свиней зарегистрировано рождение поросят с одной половой хромосомой 37, ХО, что соответствует синдрому Тернера у
человека.
Значительное количество цитогенетических исследований проведено у свиней истинных и псевдогермафродитов. При этом в разных тканях (кровь, кожа, костный мозг) обнаружены различные хромосомные наборы, в том числе нормальные, химеризм по половым хромосомам, иногда в сочетании с трисомией или моносомией - 38, ХХ/39, XXY; 37, ХО/38, ХХ/38, XY, отдельно трисомия 39, XXY.
Представляют интерес сообщения Михельмана и соавт. о различиях частот структурных и морфологических изменений хромосом в группах племенных хряков и свиноматок в сравнении с откормочными выбракованными животными. В первой группе структурные изменения обнаружены в 17,9 %, во второй — в 13,7 и в третьей —в 20,7 % метафаз. Морфологические изменения хромосом наблюдали соответственно в 0,58; 0,61 и 0,72 % метафаз. Аналогичные результаты получены и у крупного рогатого скота. Это указывает на то, что снижение плодовитости и болезни как основные причины выбраковки животных связаны не только с действием факторов внешней среды. Первичная причина нарушений функций организма может быть связана с изменениями в генотипах животных.
Овцы. Нормальный кариотип овец состоит из 54 хромосом, что впервые указано советским генетиком Живаго (1931). У  овец, как и у других видов животных, обнаружены различные виды аберраций хромосом. Так, в Новой Зеландии, в стране с развитым овцеводством, наблюдали три типа* робертсоновской транслокации — между 5-й и 26-й, 8-й и 11-й, 7-й и 25-й хромосомами, получившими название Массей I, Массей II и Массей III. Однако, как показали Бруер и Чепман, плодовитость у животных была нормальной. Это объясняется естественной выбраковкой не сбалансированных по числу хромосом гаплоидных клеток еще до вступления их в оплодотворение. У овец описаны также отдельные варианты реципрокных транслокаций, которые сопровождались пониженной плодовитостью животных.
овец, как и у других видов животных, обнаружены различные виды аберраций хромосом. Так, в Новой Зеландии, в стране с развитым овцеводством, наблюдали три типа* робертсоновской транслокации — между 5-й и 26-й, 8-й и 11-й, 7-й и 25-й хромосомами, получившими название Массей I, Массей II и Массей III. Однако, как показали Бруер и Чепман, плодовитость у животных была нормальной. Это объясняется естественной выбраковкой не сбалансированных по числу хромосом гаплоидных клеток еще до вступления их в оплодотворение. У овец описаны также отдельные варианты реципрокных транслокаций, которые сопровождались пониженной плодовитостью животных.
В Институте экспериментальной биологии Казахстана установлено, что у каракульских баранов в очень молодом и старом возрастах хромосомные аберрации в генеративной ткани встречаются значительно чаще, чем в среднем возрасте. Хромосомные аберрации были обнаружены у некоторых мертворожденных с врожденными аномалиями ягнят.
Лошади. Нормальный кариотип домашней лошади состоит из 64 хромосом. Точное число хромосом этого вида животных установили Сасаки и Макино только в 1962 г. Количество цитогене-тических исследований у лошадей пока еще невелико, поэтому установить истинную частоту аберраций хромосом не представляется возможным. Однако Виллер и Визнер (1981) считают, что по крайней мере 5—10 % всех зигот у лошадей, как и у других видов млекопитающих животных, могут содержать хромосомные аномалии. 90 % этих зигот отмирают на первой стадии беременности.
Исследования кариотипа лошадей показывают, что наиболее часто у этого вида встречаются аномалии в системе половых хромосом, которые вызывают бесплодие у животных (табл. 53).
53. Классификация гоносомальных аберраций у лошади
| Признак | Цитогенетический статус (кариотип) | Симптоматика | Этиология/патогенез |
| Дисгене- | 63, ХО с ати- | Периферическая ин- | |
| зия яични- | пичной Х-хро- | версия с последую- | |
| ков | мосомой | щей транслокацией | |
| 63, ХО | Нарушение цикла, руди- | Простая моносомия | |
| синдром | ментарные яичники, апла- | вследствие нерасхож- | |
| Тернера | зия или гипоплазия овоци- | дений | |
| тов, гипо- или анеструс, | |||
| гипоплазия гениталий, | |||
| задержка роста | |||
| 65, XXX | Простая трисомия | ||
| синдром | вследствие нерасхож- | ||
| сверхсамок | дений |
Продолжение
| Признак | Цитогенетический статус (кариотип) | Симптоматика | Этиология/патогенез |
| 64, XY/63, ХО | Переход к псевдогермафро- | Мозаичность | |
| или 64, ХХ/63, | дитарной феминизации | ||
| ХО | Л | ||
| 64, ХХ/64, Y | Обычно без клинических | Постзиготический | |
| женская химера | нарушений | химеризм | |
| Дисгенсзия | 65.XXY синдром | Гипоплазия семенников, аплазия зародышевых | Трисомия вследствие нерасхождений |
| Клайнфельтера | гормональных клеток | ||
| (высокорослость) | |||
| 64, ХХ/65, | Повышенная агрессив- | Мозаичность | |
| ХХУили64, ХХ/64, XY/65, XXY мозаика | ность, высокая'прыгучесть | ||
| Трисомия вслед- | |||
| Клайнфельтера, | ствие нерасхождения | ||
| 65, XYY син- | |||
| дром самцов | |||
| 66, XXYY; 66, | Переход к мужскому | Тетра- или пентасо- | |
| XXXY | псевдогермафродитизму | мия вследствие нерасхождения | |
| 66, XXYY; 66, | |||
| XXXY;67. | |||
| XXXYY;67, XXXXY | |||
| Комплексный | |||
| кариотип 64, ХХ/64, XY | |||
| мужской химеризм | Зиготический химеризм | ||
| Мужской псевдогер- | 64, ХХтестику-лярные | ■ Гипоплазия гонад, стерильность, аплазия заро- | Гормональное нарушение, нарушение |
| мафроди-тизм | ХХ-герма-фродиты | дышевых клеток, частично женские половые органы (тестикулярная феминиза- | во время гипотала-мической фазы дифференциации |
| ция), часто гипоплазия | |||
| пениса и крипторхизм | |||
| 64, ХУтести- | Гормональное нару- | ||
| кулярные XY- | шение, адренальное | ||
| гермафродиты | сверхпродуцирование эстрагенов, тес- | ||
| тикулярная фемини- | |||
| зация | |||
| 64, ХХ/65, | Тетрасомия вслед- | ||
| XXY или 64, | ствие нерасхождения | ||
| ХХ/64, XY/65, | Мозаичность | ||
| XXY мозаика | |||
| Клайнфельтера | |||
| 64, ХХ/65, XY | Переход к тестикулярной | Зиготический химе- | |
| тестикулярные | дисгенезии | ризм | |
| XX/XY-герма- | |||
| фродиты |
| Признак | Цитогенстичсский | Продолжение | |
| Женский псевдогер-мафроди-тизм | статус (кариотип) 64, XY овари-альные XY-гермафродиты 63, ХО/64, XY | Симптоматика Гипоплазия яичников, стерильность вследствие отсутствия зародышевых клеток. Переход к овариаль-ной дисгенезии | Эгиологкя/патогенез Гормональное нарушение во время гипоталамической фазы дифференциации |
| мозаика Тернера | Мозаичность |
Нарушения в системе половых хромосом, как показали Бруер с сотр. (1978), — общая причина стерильности у лошадей. При обследовании 7 бесплодных кобыл у 6 из них авторы обнаружили синдром Тернера (63, ХО); одна оказалась мозаиком (63, ХО/64, XX). Синдром Тернера выявлен и в наших исследованиях у двух бесплодных кобыл, используемых конноспортивными секциями Ленинградской области.
И£ структурных мутаций у лошадей описаны следующие: де-леция хромосомы второй пары у бесплодной кобылы; деления хромосомы тринадцатой пары у бесплодного жеребца американской стандартбредной породы с аномальными спермиями; моза-ицизм по Х-хромосоме с делецией длинного плеча у жеребца с низкой степенью оплодотворения; аутосомная реципрокная транслокация в гетерозиготном состоянии у жеребца с нормальной воспроизводительной системой; транслокация гоносомы и аутосомы у кобылы с такими же признаками, как при синдроме Тернера, и частичная трисомия одной аутосомы у жеребца с тяжелым клиническим эффектом; транслокация дистальной части длинного плеча Х-хромосомы на длинное плечо другой Х-хромосомы у кобылы, 64,X/t(X). Buoen с соавт. предполагают наличие изохромосом у двух лошадей. Еще одна структурная мутация выявлена в системе половых хромосом. Rayne с соавт. предполагают перицентрическую инверсию в Х-хромосоме у бесплодной, моносомной по половым хромосомам кобылы.
Птицы. В нормальном кариотипе кур, цесарок и перепелок содержится 78 хромосом, индеек и уток — 80. Установлено, что причина большей части раннего отмирания эмбрионов кур — аберрации хромосом. Ялар и Фечхаймер считают, что, как минимум, 25 % гибели эмбрионов обусловлено аномалиями в кариотипе.
У эмбрионов обнаруживали структурные нарушения — транс-локации, изохроматидный разрыв и др. Наиболее часто выявляют гетерошюидные формы аномалий кариотипа — гагагоидию, тригагоидию, трисомию по аутосомным и половым хромосомам, мозаицизм.
В экспериментах Фечхаймера показано, что гаплоидные эмбрионы кур развивались на основе андрогенеза. Триплоидные клетки в 75 % случаев были результатом нарушений при образовании второго полярного тельца. Тетраплоидные клетки формировались в основном за счет нарушения цитокинеза в митозе.
Ю. А. Эрматов и А. Ф. Яковлев установили возрастание частоты аберраций хромосом у эмбрионов, полученных от искусственного осеменения спермой, хранившейся 4 мес в замороженном состоянии. Это указывает на необходимость цитогенетичес-кого контроля при разработке технологии замораживания спермы.
Для получения оплодотворенных инкубационных яиц важное значение имеют сроки совместного содержания петухов и кур. В эксперименте Попеску и Меррита (1977) установлено резкое снижение оплодотворенности яиц после 12 дней с момента отсадки петухов от кур, увеличение общего процента эмбрионов с аномалиями хромосом. Таким образом, при длительном нахождении спермиев в половом тракте у кур происходят патологические процессы «старения», приводящие к развитию аномальных эмбрионов. Это относится и к другим видам животных. Поэтому определение момента осеменения самок — одно из условий плодотворности зачатия.
При исследовании кариотипов кур в постэмбриональный период установлены различия между популяциями и линиями по частоте хромосомных аномалий (табл. 54).
54. Хромосомные аберрации в разных линиях кур (по Блому, 1974)
| Лнння | Число исследованных несушек | Число несушек, унаследовавших одну или более хромосомных аберраций | Доля дефектных иесушек, % |
| 1-я | |||
| 2-я | |||
| 3-я | |||
| 4-я | |||
| 5-я | |||
| 6-я | |||
| 7-я | |||
| 8-я | |||
| 9-я |
По данным Фечхаймера, среди аномальных клеток у кур наиболее часто встречается гаплоидия. Цыплята мясных пород (бройлеры) имели в 7 раз больше гетероплоидных клеток, чем цыплята яйценоских пород. Причиной повышенной частоты хромосомных аномалий у кур мясного типа могут быть неправильные овуляционные периоды. Установлено, что первое мейо-тическое деление происходит у них на 2 ч раньше овуляции, а второе — при оплодотворении.
Из приведенных материалов следует вывод: цитогенетический анализ можно использовать в селекции птиц для браковки кур и петухов с хромосомными аномалиями. Это позволит повысить выводимость птицы.
Контрольные вопросы. 1. Что включают в себя понятия генетические, наслед-ственно-средовые, экзогенные аномалии? 2. Каков ход генетического анализа при простом аутосомном рецессивном типе наследования аномалии? 3. Каковы основные правила наследования аутосомно-рецессивных и аутосомно-доминантных аномалий? 4. В чем заключаются особенности наследования сцепленных с Х-хромосомой аномалий? 5. Что входит в понятие «мультифакториальное наследование» и каковы его особенности? 6. Что понимают под терминами «пенет-рантность» и «экспрессивность»? Какова их роль при наследовании аномалий? 7. Какие примеры генетических аномалий у крупного рогатого скота свиней, овец и лошадей вы знаете? 8. Как прилагается закон гомологических рядов в наследственной изменчивости для изучения генетических аномалий у животных? 9. Каково распространение аномалий хромосом у крупного рогатого скота? Какие аберрации наиболее часто встречаются у этого вида? 10. Каков спектр аберраций хромосом, обнаруженных у свиней? 11. Какие аберрации хромосом часто служат причиной бесплодия кобыл и жеребцов? 12. В чем особенности спектра аберраций хромосом птиц?








