 2014-02-09
2014-02-09 3366
3366Работа дыхательных ферментов регулируется с помощью эффекта, который получил название дыхательный контроль.
Дыхательный контроль – это прямое ингибирующее влияние электрохимического градиента на скорость движения электронов по дыхательной цепи.
При возрастании протонного градиента его ингибирующее влияние усиливается и продвижение электронов по цепи замедляется. В результате снижается выкачивание ионов Н+в межмембранное пространство, что вызывает снижение протонного градиента и ускорение электронов. Таким образом, величина электрохимического градиента остается на определенном среднем уровне.
Дополнение
При интенсивной мышечной работе количество АТФ в клетке быстро уменьшается, это вызывает накопление АДФ, являющегося субстратом для АТФ-синтазы. АТФ‑синтаза начинает работать, ионы Н+ устремляются в матрикс и величина электрохимического градиента снижается. По принципу дыхательного контроля это ускоряет движение электронов по дыхательной цепи, что имеет два очень важных последствия:
1. Возрастает выкачивание ионов Н+. Восстанавливаемый протонный градиент используется для пополнения запасов АТФ.
2. Так как источником электронов являются НАДН и ФАДН2, в соответствующих участках увеличивается их окисление. В результате снимается ингибирующее влияние НАДН на цикл лимонной кислоты и пируватдегидрогеназный комплекс. Активируются реакции катаболизма углеводов и жиров, как источника энергии.
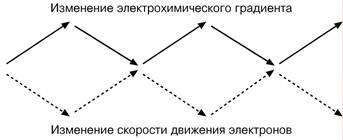 Скорость движения электронов и величина электрохимического градиента всегда изменяются в противофазе: повышение одной характеристики снижает величину другой.
Скорость движения электронов и величина электрохимического градиента всегда изменяются в противофазе: повышение одной характеристики снижает величину другой.
Коэффициент Р/О
Коэффициент Р/О – это отношение количества неорганического фосфата, включенного в молекулу АТФ АТФ-синтазой к количеству атомов кислорода, включенного в молекулу Н2О, при переносе одной пары электронов по дыхательной цепи.
Для расчета Р/О при окислении какой-либо молекулы необходимо знать:
· каждый восстановленный эквивалент (молекулы НАДН или ФАДН2) передает в цепь переноса электронов по 2 электрона.
· для восстановления кислорода в воду необходима 1 пара электронов.
· при прохождении пары электронов через всю дыхательную цепь (т.е. через I, III, IV комплексы дыхательных ферментов) ионов Н+ выкачивается столько, сколько необходимо для синтеза 3 молекул АТФ. Зная, что НАДН окисляется в I комплексе, ясно, что при его использовании образуется 3 АТФ или, по-другому, в состав АТФ включается три атома неорганического фосфата.
· при прохождении пары электронов через III и IV комплексы дыхательных ферментов ионов Н+ выкачивается столько, сколько необходимо для синтеза 2 молекул АТФ. Зная, что ФАДН2 окисляется в III комплексе, ясно, что при его использовании образуется 2 АТФ или, по-другому, в состав АТФ включается два атома неорганического фосфата.
Таким образом, коэффициент Р/О для НАДН равен 3, коэффициент Р/О для ФАДН2 равен 2.
Расчет энергетической ценности
и коэффициента Р/О
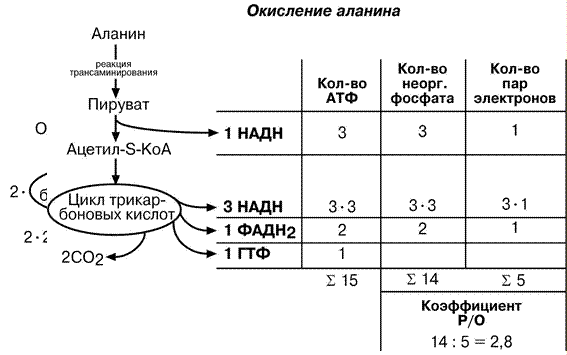 |
При расчете энергетической ценности, т.е. количества АТФ, образующейся при окислении аминокислот и коэффициента Р/О необходимо представлять себе весь путь аминокислоты до полного окисления ее углеродных атомов до СО2. При этом необходимо учитывать число атомов углерода в молекуле.
Например, аспартат содержит 4 атома углерода и поэтому его остатку необходимо пройти 2 оборота ЦТК, прежде чем они выделятся в виде СО2. Подсчитывая число восстановленных НАДН, ФАДН2 и ГТФ, образуемых в двух оборотах ЦТК, определяем сумму АТФ – 24 молекулы. При расчете коэффициента Р/О учитываем только фосфат, включенный в АТФ ферментом АТФ-синтазой, т.е. в процессе окислительного фосфорилирования. Это значит, что фосфат, входящий в ГТФ, не учитывается! И, наконец, помним, что каждая молекула восстановленного эквивалента (т.е. НАДН или ФАДН2) передает на дыхательную цепь по одной паре электронов, которые проходят разное расстояние: от НАДН – три комплекса ферментов, от ФАДН2 – 2 комплекса ферментов, но в любом случае восстанавливают до воды по одному атому кислорода.
Схожим образом рассчитываем требуемые значения суммы АТФ и коэффициента Р/О для аланина и глутамата.

Два атома углерода аланина попадают в ЦТК после того, как безазотистый остаток аланина – пируват – окислится в пируват-дегидрогеназном комплексе с образованием НАДН. Образованный ацетил-S-КоА входит в ЦТК и полностью окисляется в одном его обороте. В результате в дыхательную цепь направляется 4 молекулы НАДН и 1 молекула ФАДН2.
В случае с глутаматом надо учесть, что a-кетоглутарат, образуемый из глутаминовой кислоты, является метаболитом цикла Кребса. Он сразу превращается в сукцинил-S-КоА и далее по реакциям ЦТК до оксалоацетата. Для окисления оксалоацетата требуется два оборота ЦТК (см выше).

