 2014-02-09
2014-02-09 6800
68001. Пигментный аппарат растений. Фотосистемы.
2. Световая стадия фотосинтеза. Фотофосфорилирование.
3. Темновая стадия фотосинтеза. Цикл Кальвина. С4-путь. САМ-метаболизм.
4. Фотодыхание.
5. Экология фотосинтеза.
6. Значение фотосинтезирующих организмов для биосферы
1. Фотосинтез – это процесс синтеза органических (глюкозы) соединений из неорганических (СО2 и Н2О) за счет поглощенной энергии света. Фотосинтез осуществляют высшие растения, водоросли и некоторые бактерии. Он является основным процессом в энергетике биосферы.
Пигменты- важнейший компонент аппарата фотосинтеза. Впервые разделены пигменты зеленого листа были в 1903 году М.С.Цветом с помощью адсорбционной хроматографии.
Пигменты пластид относятся к трем классам веществ: хлорофиллам, фикобилинам и каротиноидам.
Хлорофиллы. Основу молекулы хлорофилла, как и гемагемоглобина составляет порфириновое кольцо. Элементарный состав молекулы хл а – С55Н72О5N4Mg и хл b - С55Н70О6N4Mg. Структура была расшифрована немецким биохимиком Г.Фишером: хлорофилл – это сложный эфир дикарбоновой кислоты хлорофиллина, у которой одна карбоксильная группа этерифицирована остатком метилового спирта, а другая - остатком одноатомного непредельного спирта фитола
Рис. 1Структурная формула хлорофилла: 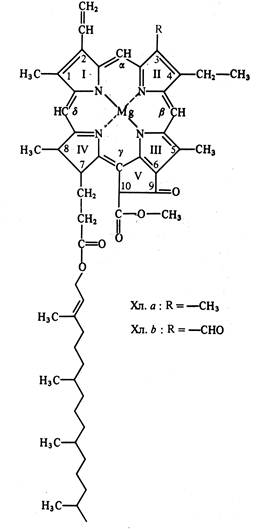
Порфириновое ядро образовано 4 пиррольными кольцам, атомы азота которых соеденины координационными связями с атомом магния, а в целом структура представляет собой систему чередующихся сопряженных двойных и одинарных связей с 18 делокализоанными p- электронами.
У высших растений и водорослей обнаружены хл a,b,c,d. Причем хл а есть у всех фотосинтетиков, хл b- у высших растений, зеленых водорослей и эвгленовых, хл c- у бурых и диатомовых водорослей, хл d- у красных водорослей, у бактерий – бактериохлорофиллы.
Синтезируется хлорофилл в растениях в хлоропластах из С5-дикарбоновых кислот (н-р, глутаминовой). Хлорофиллы хорошо растворимы в этиловом спирте, бензоле, ацетоне, этиловом эфире, плохо растворимы в петролейном эфире и нерастворимы в спирте.
Раствор хлорофилла а имеет сине-зеленый цвет, хлорофилла b – желто-зеленый. Максимумы поглощения хлорофиллов лежат в красной и синей части спектра: 660-663 и 428-430; и 642-644 и 452-455 нм соответственно. Хлорофиллы совсем не поглощают оранжевый и инфракрасные лучи. Поглощение в сине-фиолетовой части спектра обусловлено системой коньюгированный одинарных и двойных связей порфирирового кольца молекулы хлорофилла, а в красной области – присутствием магния.
Растворы хлорофиллов в полярных растворителях обладают яркой рубиново-красной флуоресценцией (люминисценцией) и фосфоресценцией (длительному послесвечению) в инфракрасной области. Механизм этих явлений заключается в том, что электроны пигментов за счет энергии света из основного синглетного энергетического состояния (So) на 10-13 – 10-9 с переходят в возбужденное синглетное состояние (S*) или на 10-4 с - в триплетное (Т*), при обращении спина. Возврат молекулы в основное состояние сопровождается излучением кванта света с большей длиной волны (правило Стокса), то есть красного в первом случае и инфракрасного во втором.
Если же хлорофилл находится в составе светопоглощающего комплекса, то есть закреплен с помощью фитола в мембране тилакоида хлоропласта, то энергия возбужденного состояния используется на фотохимические реакции фотосинтеза.
Фикобилины водорослей по структуре относятся к группе желчных пигментов. Это тетрапирролы с открытой цепью, имеющие систему коньюгированных двойных и одинарных связей, обычно связанные с глобулиновыми белками. Бывают: 1) фикоэритрины- красные с максимумами поглощения 498-568 нм; 2) фикоцианины – сине-голубые, 585-630 нм и 3) аллофикоцианины - синие, 585-650 нм.
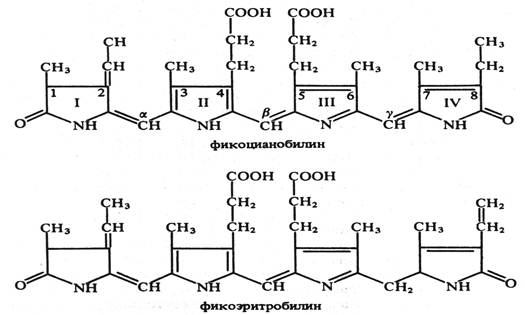
Рис. 2 Структурные формулы фикобилинов
Значение фикобиллинов – выполняют роль дополнительных пигментов, так как максимумы их поглощения лежат в оранжевой, желтой и зеленой частях спектра. Значение этих лучей возрастает с глубиной, так вода обладает способностью поглощать в первую очередь длинноволновые лучи.
Каротиноиды – жирорастворимые пигменты желтого, оранжевого, красного цвета – присутствуют в хлоропластах всех растений. К ним относят 1) оранжевые и красные каротины (С40Н56); 2) желтые ксантофилы (С40Н56О2 и С40Н56О4). Каротины и ксантофилы хорошо раствормы в хлорофирме, бензоле, ацетоне. Каротины легко растворимы в петролейном и диэтиловом эфире, но почти не растворимы в метаноле и этаноле. Ксантофиллы хорошо растворимы в спиртах и значительно хуже в петролейном эфире.
Все каротноиды – полиеновые соединения, состоящие из восьми остатков изопрена, которые образуют цепь коньюгированных двойных связей обычно с циклами на концах.
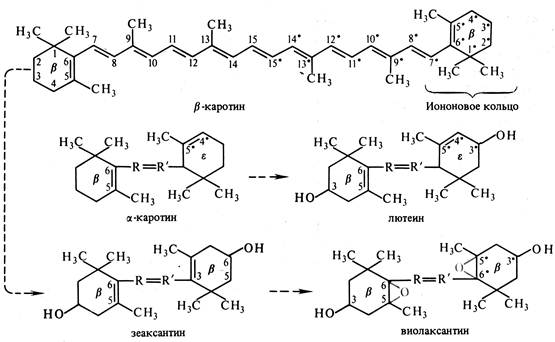
Рис.3 Структурные формулы фикобилинов
К этой группе пигментов относят лютеин, зеаксантин кукурузы, виолаксантин и фукоксантин бурых водорослей. Синтезируются каротиноиды из ацетил-Со А через мевалоновую кислоту.
Спектры поглощения каротиноидов характеризуются двумя полосами в фиолетово-синей и синей области от 400 до 500 нм и определяется системой коньюгированных двойных связей.
Каротиноиды – обязательные компоненты пигментных систем всех фотосинтезирующих организмов. Главные функции: 1)участие в поглощении света в качестве дополнительных пигментов; 2) защита молекул хлорофиллов от необратимого фотоокисления.
Впервые идею о том, что в хлоропластах существует как минимум две пигментные системы высказал Р.Эмерсон в 1957. Оказалось, что суммарный эффект (квантовый выход) фотосинтеза был выше при освещении хлореллы одновременно коротковолновым (650 нм) и длинноволновым (700 нм) светом – эффект усиления Эмерсона.
Позже удалось выделить и изучить комплексы фотосистемы I и фотосистемы II (ФС I, ФС II).
Каждая ФС состоит из антенного или светособирающего комплекса (ССК) из 200-400 молекул пигментов и реакционного центра.
В состав ФС I в качестве реакционного центра входит димер хлорофилла а П700., а антенный комплекс – это хромопротеины, содержащие 110 молекул хлорофиллов а с максимумами поглощения 680-695 нм на каждый П700, 80-100 молекул хлорофилла b и b-каротин.
В состав ФС II в качестве реакционного центра входит хлорофилл а П680., а антенный комплекс – это хромопротеины, содержащие 40 молекул хлорофиллов а с максимумами поглощения 670-683 нм на каждый П680, 30 молекул хлорофилла b и b -каротин.
2. На световой фазе фотосинтеза происходит поглощение света молекулами пигментов и трансформация этой энергии в химическую энергию АТР и восстановленного NADPH. Все эти процессы осуществляются в фотохимически активных мембранах хлоропластов и представляют собой сложную систему фотофизических и фотохимических реакций.
Фотофизический этап. Роль пигментных антенных комплексов состоит в том, чтобы собирать и передавать энергию квантов света, которую каждая молекула пигмента поглощает не чаще одного раза за 0,1с, на молекулы реакционных центров, поддерживая его таким образом все время в возбужденном состоянии, «без простоя». Передача энергии происходит по принципу индуктивного резонанса, когда электрическое поле вокруг возбужденной молекулы с определенной частотой колебаний индуцирует осцилляцию диполя (электрон – ядро) соседней (расстояние не более 10 нм) молекулы и ее переход в возбужденное состояние.
Таким образом, в антенных комплексах перенос энергии осуществляется в ряду: каротин (400-550 нм) хлорофилл a (650нм) хлорофилл b (660-675 нм) П680.Скорость резонансного переноса энергии от молекулы к молекуле 10-10 – 10-9 с.
Миграция энергии к реакционному центру завершает фотофизический этап фотосинтеза, в результате которого энергия квантов света была переведена в энергию возбужденного состояния молекул реакционных центров.
Фотохимический этап. Молекулы пигментов реакционных центров способны к первичному фотохимическому разделению зарядов и связано это с транспортом электронов. В РЦ ФС11 первичным донором электронов служит П680, а первичным акцептором – белок феофетин (Фф). В РЦ ФС1 – донор – П700, акцептор – мономерная форма хлорофилла a695 (А1). Вторичными акцепторами являются:
- В ФС 1 – железосерные белки А2 и Ав и ферредоксин(Фд);
- В ФС 11 – пластохиноны (QA, QB и PQ).
Нециклический и циклический транспорт электронов (Z-схема).
Впервые принцип Z- схемы последовательных реакций транспорта электронов была разработана Р.Хиллом и Ф.Бендаллом в 1960 году и является общепризнанной.
В ФС11 димер П680, поглотив энергию 2 квантов переходит в возбужденное сосотояние и отдает 2 электрона Фф. От Фф электроны передаются на пластохиноны QA, QB, затем на пул липидорастворимых молекул PQ, которые переносят электроны и протоны через липидную фазу мембраны на железосерный белок Риске FeSR и цитохром f ( гемопротеин ) цитохромного комплекса b 6- f, восстанавливая Сu-содержащий белок пластоцианин (Пц).
Вакантные места («дырки») в П680 заполняются двумя электронами из содержащего марганец переносчика электронов Z, который в свою очередь восстанавливается с участием системы S. Окисленный белковый комплекс S связывает воду (фотолиз) и восстанавливается за счет электронов воды. Для осуществления этих реакций в белковом комплексе S необходимы Mn и Cl-.
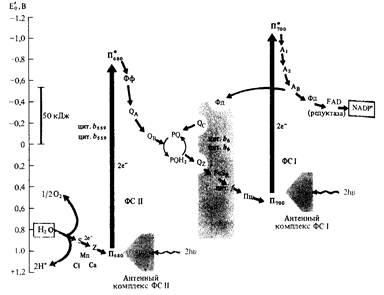 |
При возбуждении П700 в реакционном центре ФС1 2 электрона захватываются мономерной формой хлорофилла а (А1) и передаются последовательно на А2, Ав и Фд, с которого электроны с помощью NADP-оксидоредуктазы с FAD в качестве кофактора идут на восстановление NADP+.
На вакантные места в П700 переходят электроны с Пц и нециклическая цепь переноса электронов таким образом замыкается.
Наряду с нециклическим в мембранах хлоропластов функционирует циклический транспорт электронов, включающий в себя только ФС1 и комплекс цитохромов b6-f. В этом случае NADP+ не восстанавливается и освобождающаяся энергия используется для фосфорилирования АDP.
Фотофосфорилирование.
Энергия, освобождающаяся при движении электронов от П680 (Е0=-0,8В) до П700 (Е0=+0,4В), что составляет >50 кДж, используется для синтеза АТР из АDP и фосфора нерганического.
Механизм фотофосфорилирования, сопряженного с деятельностью электронтранспортной цепи, обьясняет хемиосмотическая теория П.Митчелла. Сущность теории состоит в следующем. Цепь переносчиков электронов и протонов, действующая в соответствии с окислительно-восстановительным градиентом, перешнуровывает мембрану таким образом, что трансмембранный перенос е- и Н+ в одну сторону чередуется с переносом в обратную сторону только е-. В результате функционирования такого механизма (Н+-помпы) по одну сторону мембраны накапливается избыток Н+ и возникает электрохимический (т.е. электрический и концентрационный градиент ионов Н+), который служит формой запасания энергии. Обратный пассивный ток ионов Н+ через протонный канал Н+-АТР-азы сопровождается образованием высокоэнергетической связи АТР.
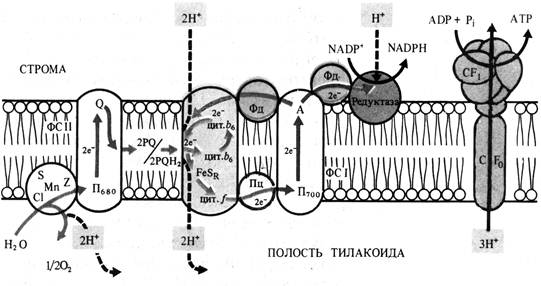
Рис. 4. Локализация электрон- и протонтранспортных реакций в тилакоидной мембране хлоропласта
В результате фотохимических реакций в хлоропластах создается необходимый уровень АТР и NADPH, которые необходимы для функционирования темновой стадии фотосинтеза, где СО2 восстанавливается до углевода.
3. Существуют различные пути восстановления СО2. Рассмотрим каждый из них.
1) С3-фотосинтез или цикл Кальвина.
Этот способ ассимиляции присущ всем растениям и впервые был описан Американским биохимиком М.Кальвином в 1956г.С помощью меченого 14СО2 удалось обнаружить, что первичным продуктом фотосинтеза является С3 соединение фосфоглицериновая кислота (ФГК) – отсюда и название пути, а первичным акцептором СО2 – рибулозо-1,5-дифосфат (РБФ).
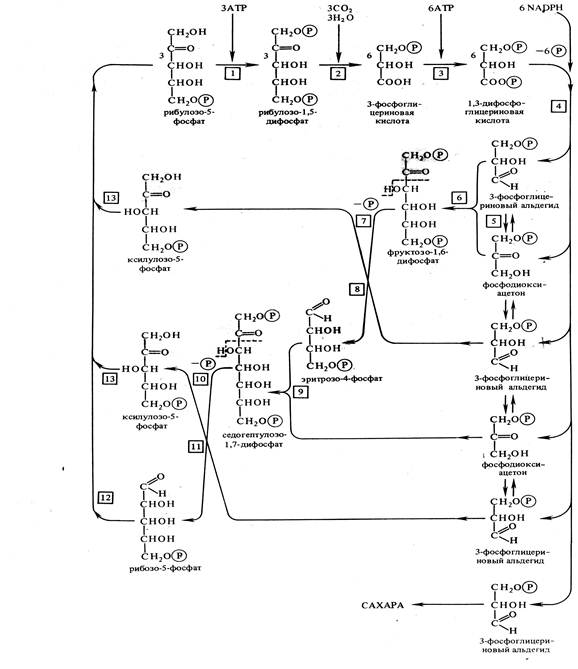
Рис. 5. Цикл Кальвина (С3-путь фотосинтеза).
1 – фосфорибулокиназа, 2 – рибулозодифосфаткарбоксилаза, 3 - фофсфоглицераткиназа, 4 – триозофосфатдегидрогеназа, 5 – триозофосфатизомераза, 6 – альдолаза, 7 – фосфатаза, 8 – транскетолаза, 9 – альдолаза, 10 – фосфатаза, 11 – транскетолаза, 12 – рибозофосфатизомераза, 13 – фосфокетопентоэпимераза
Весь процесс состоит из трех этапов:
-Карбоксилирование. Молекула рибулозо-5-фосфата фосфорилируется с участием АТР, а к образовавшемуся РДФ с помощью РДФ-карбоксилазы присоединяется поступивший в мезофилл через устьица СО2.Полученный продукт тут же распадается на две триозы: две молекулы 3-ФГК.
- Фаза восстановления. 3-ФГК восстанавливается до 3-ФГА (альдегида) в два этапа: сначала происходит фосфорилирование 3-ФГК с помощью АТР до 1,3-ФГК, а затем ее восстановление до 1,3-ФГА с помощью NADPH и дегидрогеназы фосфоглицеринового альдегида.
- Фаза регенерации первичного акцептора СО2 и синтеза конечного продукта фотосинтеза. При фиксации 3 молекул СО2 из шести образовавшихся 3-ФГА пять используются для регенерации рибулозо-5-фосфата, а один – для синтеза глюкозы. Регенерация осуществляется с помощью ферментов изомераз, транскетолаз и альдолаз через промежуточные С4, С6, С7-соединения.
Из оставшейся неиспользованной шестой молекулы 3-ФГА при повторении цикла синтезируется фруктозо-1,6-дифосфат, из которого могут образовываться глюкоза, сахароза и крахмал.
2) С4-путь фотосинтеза (Цикл Хетча и Слэка).
К группе растений с С4-путем фотосинтеза относятся сахарный тростник, кукуруза, сорго и др.Листья этих растений содержат два разных типа хлоропластов: хлоропласты обычного вида – в клетках мезофилла и большое количество крупных хлоропластов в клетка, окружающих проводящие пучки (обкладка).
СО2 попадает в цитоплазму клеток мезофилла, где при участии ФЕП-карбоксилазы вступает в реакцию с фосфоенолпируватом (ФЕП), образуя щавелевоуксусную кислоту (оксалоацетат). Затем уже в хлоропластах оксалоацетат восстанавливается до яблочной кислоты (малата) за счет NADPH, образующегося в ходе световой стадии фотосинтеза. Затем малат переносится в хлоропласты клетки обкладки сосудистого пучка, где он декарбоксилируется малаьдегидрогеназой декарбоксилирующей до пирувата и СО2. СО2 вступает в цикл Кальвина, а пируват возвращается в хлоропласты клеток мезофилла.

Рис. 6. Цикл Хэтча и Слэка (С4-путь фотосинтеза). КС – клеточная стенка.
Такая компартментализация процесс фиксации и использования СО2 позволяет растениям с С4-путем осуществлять фотосинтез даже при закрытых устьицах (в засушливы или засоленных местах обитания), так как хлоропласты клеток обкладки используют малат, образовавшийся ранее, как донор СО2.С4-растения могут также использовать СО2, возникающий при фотодыхании.
3) САМ-метаболизм (по типу толстянковых)
У суккулентов наблюдается суточный цикл метаболизма С4-кислот с образованием яблочной кислоты ночью. Этот тип фотосинтеза называют САМ-метаболизм (crassulacean acid metabolism).Устьица этих растений днем обычно закрыты, что предотвращает потерю воды, и открываются ночью. СО2 поступает в листья, где взаимодействует с ФЕП, образуя ЩУК (оксалоацетат), который восстанавливается до яблочной кислоты. Она накапливается в вакуолях клеток листа, что приводит к закислению клеточного сока в ночное время.
Днем в условиях высокой температуры, когда устьица закыты, малат транспортируется из вакуолей в цитоплазму и там декарбоксилируется при участии малатдегидрогеназы декарбоксилирующей (малик-энзима) с образованием СО2 и пирувата. СО2 поступает в хлоропласты и включается в них в цикл Кальвина, участвуя в синтезе сахаров.
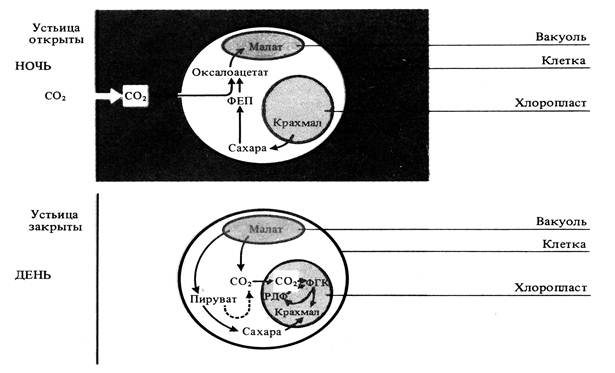
Рис. 7. Метаболизм органических кислот при фотосинтезе по типу толстянковых (САМ-метаболизм).
4. Фотодыхание – это активируемый светом процесс высвобождения СО2 и поглощения О2, т.е. процесс обратный фотосинтезу. Этот процесс усиливается при низком содержании СО2 и высоких концентрациях О2 или при повышении температуры. В этих условиях РДФ-карбоксилаза в хлоропластах может функционировать как оксигеназа, катализируя окислительное расщепление Рибулозо-1,5-дифосфата на 3-ФГК и 2-фосфогликолевую кислоту, которая дефосфорилируется в гликолевую кислоту.
Гликолат из хлоропластов поступает в пероксисому и там окисляется до глиоксилата. Возникающая перекись водорода устраняется каталазой. Глиоксилат аминируется, превращаясь в глицин и транспортируется в митохондрию.
В митохондрии из двух молекул глицина образуется серин и освобождается СО2. Серин может снова поступать в пероксисому, где трансаминируется с пируватом. Образовавшийся гидроксипируват восстанавливается до глицерата, который может вновь поступать в хлоропласты и включаться в цикл Кальвина.
У С4-растений высвобождающийся при фотодыхании СО2 перехватывается в клетках мезофилла, где из него образуются оксалоацетат и малат. 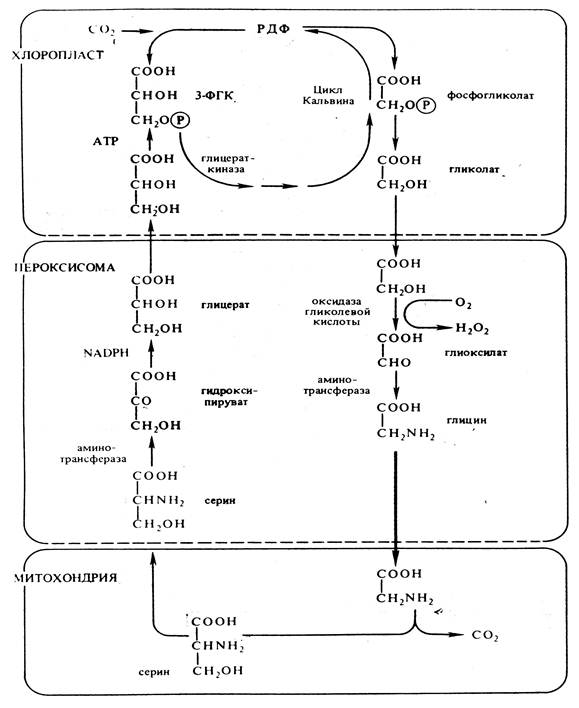
Рис. 8. Фотодыхание.
5. Фотосинтез осуществляется в полуавтономных органеллах – хлоропластах. Однако он в значительной мере контролируется процессами, происходящими в растении, и факторами внешней среды
Отток ассимилятов. Накопление фотоассимилятов в хлоропластах и в околопластидном пространстве приводит к ингибированию ферментов, участвующих в фотосинтезе.
Содержание хлорофилла. С увеличением содержания в клетке хлорофилла увеличивается интенсивность фотосинтеза.
Возраст листа и растения. В ходе роста листа интенсивность фотосинтеза увеличивается. После окончания роста листа она постепенно снижается. У многих однолетних растений интенсивность фотосинтеза достигает максимума в фазу бутонизации и цветения, а затем снижается.
Свет. Имеется нижний порог освещенности, при котором растения начинают фотосинтезировать. Затем зависимость интенсивности фотосинтеза от освещенности имеет логарифмический характер с последующим выходом на плато. Угол наклона кривой зависимости интенсивности фотосинтеза от освещенности зависит от влияния других факторов. Так, у светолюбивых растений она выходит на плато при значительно более высокой освещенности, чем у теневыносливых растений. Уровень освещения, при котором поглощение СО2 в ходе фотосинтеза равно выделению СО2 в процессе дыхания, называется компенсационным пунктом.
Важен и спектральный состав света. При освещении красным светом образуются преимущественно углеводы, синим - амино- и органические кислоты.
Температура. При низкой освещенности фотосинтез идет с одинаковой скоростью при 15 и 25оС. Это объясняется тем, что при низкой освещенности интенсивность фотосинтеза зависит от скорости световых реакций. При высокой освещенности интенсивность фотосинтеза лимитируется скоростью темновых реакций и Q10 примерно равен 2. Для большинства растения С3-типа оптимальная температура 20-25оС, для растений С4-типа она равна 25-40оС. При температуре выше оптимальной интенсивность фотосинтеза снижается из-за инактивации хлоропластов и закрытия устьиц.
Содержание СО2 в воздухе. Повышение содержания СО2 с 0,03 % до 0,3 % вызывает увеличение интенсивности фотосинтеза. Дальнейшее возрастание концентрации СО2 до 1 % не сказывается на фотосинтезе, но более высокий уровень СО2 в воздухе приводит к депрессии фотосинтеза. Высокие концентрации СО2 особенно неблагоприятны при высокой освещенности, так как происходит ингибирование темновых реакций. Влияние содержания углекислого газа на фотосинтез зависит от вида растения.
Снабжение водой. При большом водном дефиците интенсивность фотосинтеза снижается из-за закрытия устьиц, что уменьшает поступление СО2 в листья, снижает транспирацию и приводит к повышению температуры листа. Кроме того, обезвоживание изменяет конформацию и, следовательно, активность ферментов.
Содержание кислорода в воздухе, в среднем, равно 21 %. Повышение концентрации или отсутствие кислорода для фотосинтеза неблагоприятны. Кислород снижает активность рибулозодифосфаткарбоксилазы.
Минеральное питание. Исключение любого элемента минерального питания отрицательно сказывается на фотосинтезе. Особенно важны такие элементы как фосфор, магний, железо, марганец, медь, калий и азот. На всех этапах фотосинтеза участвуют фосфорилированные соединения. Калий активирует процессы фосфорилирования и участвует в открывании устьиц. Магний входит в состав хлорофиллов, активирует реакции карбоксилирования и восстановления НАДФ. Железо необходимо для синтеза хлорофиллов. Марганец участвует в фоторазложении воды. Медь входит в состав пластоцианина. Азот необходим для формирования хлоропластов и образования пигментов.
6. Выделяют пять основных аспектов планетарной роли фотосинтезирующих организмов.
Накопление органической массы. В процессе фотосинтеза наземные растения образуют до 170 млрд. тонн, а растения мирового океана – до 70 млрд. тонн биомассы в год в пересчете на сухое вещество, используемой гетеротрофными организмами.
Обеспечение постоянства содержания СО2 в воздухе. Связывание СО2 в ходе фотосинтеза в значительной мере компенсирует его выделение в результате других процессов (дыхание, брожение, деятельность вулканов, производственная деятельность человечества).
Препятствие развитию парникового эффекта. Часть солнечного света отражается от поверхности Земли в виде тепловых инфракрасных лучей. СО2 поглощает инфракрасное излучение и тем самым сохраняет тепло на Земле. Повышение содержания СО2 в атмосфере может способствовать увеличению температуры, то есть создавать парниковый эффект. Это приведет к затоплению прибрежных зон из-за поднятия уровня мирового океана в результате таяния ледников в горах и на полюсах. Однако высокое содержание СО2 в воздухе активирует фотосинтез и, следовательно, концентрация СО2 в воздухе опять уменьшится.
Накопление кислорода в атмосфере. Первоначально в атмосфере Земли кислорода было очень мало. Сейчас его содержание составляет 21 % по объему воздуха. В основном, этот кислород является продуктом фотосинтеза. Ежегодно растения и другие фотосинтезирующие организмы поставляют в атмосферу примерно 120 млрд. тонн кислорода.
Озоновый экран. Озон (О3) образуется в результате фотодиссоциации молекул кислорода под действием солнечной радиации на высоте около 25 км. Озон задерживает большую часть ультрафиолетовых лучей, губительных для всего живого.

