 2015-02-24
2015-02-24 10132
101321. История развития учения о минеральном питании растений.
Первые труды появляются в средние века. Среди них назовем «Научный трактат о различных почвах (солях) и сельском хозяйстве» французского естествоиспытателя Палисси. В этом сочинении впервые почва рассматривается как источник питания растений минеральными солями, высказывается мысль о необходимости возврата в почву минеральных веществ в виде удобрений.
Ван-Гельмонт. Им был поставлен эксперимент с растениями, на основании которого был сделан ошибочный вывод о питании растений водой. Ива и горшок с землей (Водная теория, была ошибочна)
Глаубер выдвинул гипотезу, согласно которой основой роста растений является селитра(«соль плодородия»), получаемую из смеси навоза с землей. Глаубер предположил, что она образуется из пищи животных. Он советовал вносить селитру под виноградники, смачивать раствором селитры высеваемое зерно.
Лавуазье установил закон сохранения вещества, определил состав воздуха и процесс образования углекислого газа, сделал ряд других важных открытий. Вместе с тем он занимался и вопросами, относящимися к агрономической химии. Он писал: «Растения черпают материалы, необходимые для своей организации, в воздухе, который их окружает, в воде, вообще в минеральном царстве.».
Пристли проделал замечательный опыт. Он взял стеклянный колпак, изолировал под ним мышь и горящую свечу, потом поместил мяту. Вывод: под действием зеленых растений воздух становится пригодным для дыхания и горения.
Ингенгаус обнаружил, что только растения, причем только на свету, поглощают из воздуха углекислоту, а выделяют при этом кислород. Сами растения непрерывно дышат, но на свету значительно больше выделяют кислорода, а при отсутствии света сами используют некоторую часть O2.
Сенебье и Соссюр представили экспериментальные доказательства минерального корневого и воздушного питания растений. Ученые показали, что в усвоении углекислого газа участвует «зеленый крахмал», то есть хлорофилл. Ученые показали, что под влиянием солнечной радиации листья испаряют влагу, способствуя этим доступу новых порций «соков» из почвы и корней в надземные органы растений. Под действием света они поглощают углекислый газ и выделяют кислород. Корни из почвы доставляют растениям лишь малую часть пищи, но эти минеральные вещества им совершенно необходимы.
Тэер был сторонником гумусовой теории питания растений. Согласно этой теории, перегной непосредственно поглощается корнями и служит основной пищей растениям. Минеральные вещества эта теория ошибочно рассматривала лишь как вспомогательные, содействующие усвоению гумуса.
Либих показал, что все минеральные соединения растения поглощают из почвы. Поэтому для восстановления почвенного плодородия эти вещества в почву необходимо возвращать, «закон возврата»; «закон минимума»: уровень урожая зависит от количества того минерального вещества, которое находится в минимуме. При этом увеличение содержания других минеральных веществ не обеспечивает увеличения урожая.
Буссенго поставил ряд точных экспериментов, в результате которых доказал, что растения не используют азот, содержащийся в воздухе. Установив, что клевер и люцерна обогащают почву азотом, Буссенго предположил, что эти растения поглощают азот из воздуха.
2. Содержание минеральных элементов в растениях. Классификация минеральных элементов: макро- и микроэлементы.
Зольные элементы сосредоточены в тех органах и клетках, уровень жизнедеятельности которых достаточно высок. Как правило, чем богаче почва и суше климат, тем выше содержание золы в растении. Необходимые элементы. Это те, без которых организм не может завершить свой жизненный цикл. Ю. Сакс и И. Кноп установили, что для жизнедеятельности растения, кроме С, О, Н, необходимы следующие 7 элементов: N, Р, S, К, Са, Mg, Fe. Позже была показана необходимость для растений в еще шести элементов: бора, марганца, цинка, меди, молибдена и хлора. Для некоторых растений характерны еще и другие элементы (например натрий). Все необходимые элементы корневого питания подчиняются основным правилам Либиха. Все необходимые для жизни растений элементы в зависимости от их количественного содержания в растении принято разделять на макроэлементы (содержание более 0,01%) — N, Р, S, К, Са, Mg, Fe и микроэлементы (содержание менее 0,01%) — Mn, Си, Zn, В, Mo, О
Общие значения питательных элементов: 1) входят в состав биологически важных органических веществ; 2) участвуют в создании определенной ионной концентрации, стабилизации макромолекул и коллоидных частиц (электрохимическая роль); 3) участвуют в каталитических реакциях, входя в состав или активируя отдельные ферменты.
3. Макроэлементы – K, Ca, Mg, S, P, их физиологическая роль.
Сера. Сера усваивается растениями только в окисленной форме—в виде аниона SCV. В растении основная масса аниона сульфата восстанавливается до —SH и —S—S— групп. Входит в состав трех аминокислот — цистина, цистеина и метионина, витаминов, коферментов (биотин, тиамин, коэнзим А, глютатион, липоевая кислота). Участвует в белковом обмене, аэробной фазе дыхания, синтезе жиров, в образовании макроэргической связи, в метаболизме и др.
Часть серы находится а клеточном соке в виде иона сульфата. При отмирании корня в условиях, когда ему не хватает кислорода, серосодержащие соединения распадаются с образованием сероводорода, который ядовит для корня. При недостатке серы, так же как и при недостатке азота, начинается разрушение хлорофилла, но первыми испытывают недостаток серы верхние листья.
Фосфор. Фосфор усваивается растениями в окисленной форме в виде солей фосфорной кислоты. Входит в состав нуклеиновых кислот (ДНК и РНК), нуклеотидов (АТФ, НАД, НАДФ), нуклеопротеидов, витаминов, фосфолипидов (мембраны) и др. Участвует в процессах гликолиза и аэробного дыхания. Освобождающаяся в этих процессах энергия накапливается в виде богатых энергией фосфатных связей. Принимает участие в фотосинтезе. При отсутствии фосфора а начале жизни и при последующей подкормке растения фосфорными солями листья растений некоторое время страдают из-за усиленного поступления фосфора и нарушенного в связи с этим азотного обмена.
Калий, кальций и магний усваиваются из любых растворимый солей, анионы которых не обладают токсическим действием или связанные с каким-нибудь нерастворимым веществом, обладающим кислотными свойствами. Они (K, Ca, Mg) необходимы для питания.
Роль: адсорбируясь на поверхности коллоидных частиц протоплазмы, они создают вокруг них определенные электростатические силы, они определяют оводненность и водоудерживающую силу коллоидов.
Калий. Калий в растениях содержится в больших кол-ах, особенно в их вегетативных частях. Калий оказывает большое влияние на коллоиды плазмы, он повышает их гидрофильность (разжижает плазму). Калий является также катализатором ряда синтетических процессов: как правило, он катализирует синтез высокомолекулярных веществ из более простых, способствует синтезу сахарозы, крахмала, жиров, белков, При недостатке калия процессы синтеза нарушаются, и в растении скапливаются глюкоза, аминокислоты и продукты распада других высокомолекулярных соединений. При недостатке калия на нижних листьях появляется краевой запал — края листовой пластинки отмирают, листья приобретают характерную куполообразную форму, на листьях появляются некрозы.
Кальций. Часть кальция находится в клеточном соке. Этот кальций не принимает активного участия в процессах обмена веществ, он обеспечивает нейтрализацию избыточно образующихся органических кислот. Часть кальция сосредоточена в плазме—здесь кальций понижает гидрофильность плазменных коллоидов, повышает их вязкость. Кальций входит в состав ядерного вещества, играет роль в процессах деления клетки. Участвует в образовании клеточных оболочек, формировании стенок корневых волосков. При отсутствии кальция быстро поражаются точки роста надземные частей и корня, так как кальций не передвигается из старых частей растения к молодым. Корни ослизняются, рост их почти прекращается или идет ненормально.
Магний. Входит в состав хлорофилла. Магний важен для дыхательного обмена, он катализирует ряд реакций образования фосфатных связей и их переноса. Так как богатые энергией фосфатные связи участвуют в самых различных синтезах, то без магния эти процессы не идут. При недостатке магния разрушается молекула хлорофилла, причем жилки листьев остаются зелеными, а участки тканей, расположенные между жилками, бледнеют. Это явление называется пятнистым хлорозом.
Железо. Поглощается в виде растворенных солей, комплексных и органических соединений. В растительных тканях железо частично переходит в органические соединения. Находясь в составе ферментов, он принимает активное участие в окислительно-восстановительных процессах. Железо входит в состав дыхательных ферментов (цитохрома, цитохромоксидазы, каталазы и пероксидазы). Принимает участие в образовании хлорофилла. При недостатке железа развивается хлороз. При недостатке железа изменяется окраска молодых листьев (желтеют), фотосинтез; рост растений замедляется.
4. Физиологическая роль азота в жизни растений. Источники азотного питания высших растений. Фиксация молекулярного азота.
Азот хорошо усваивается растением из солей азотной кислоты и аммония. Он является главным элементом корневого питания, так как входит в состав белков всех живых клеток. Азот является составной частью нуклеиновых кислот, входящих в состав ядра и являющихся носителями наследственности. Является частью хлорофилла, входит в состав ферментов, которые регулируют реакции обмена веществ, и ряда витаминов. Все формы азота растении превращаются в аммиачные соединения, которые, вступая в реакцию с органическими кислотами, образуют аминокислоты и амиды — аспарагин и глютамин. Аммиачный азот обычно не скапливается в растении в значительных количествах. Это наблюдается только при недостатке углеводов; в этих условиях растение не может его переработать в безвредные органические вещества — аспарагин и глютамин. Избыток аммиака в тканях зачастую приводит к их повреждению. Если растение испытывает недостаток в азоте, то это прежде всего сказывается на темпе роста. Новые побеги почти не образуются, размеры листьев уменьшаются. При отсутствии азота в старых листьях хлорофилл разрушается. Биологическая фиксация азота атмосферы имеет важное значение, благодаря ей азот переходит в формы, которые могут использовать все растительные, а через них и животные организмы.Организмы, способные к усвоению азота воздуха, можно разделить на группы: 1) симбиотические азотфиксаторы — микроорганизмы, которые усваивают азот атмосферы, только находясь в симбиозе с высшим растением; 2) не симбиотические азотфиксаторы — микроорганизмы, свободно живущие в почве и усваивающие азот воздуха; 3) ассоциативные азотфиксаторы — микроорганизмы, обитающие на поверхности корневой системы злаков, т. е. живущие в ассоциации с высшими растениями. Важное значение имеют симбиотические азотфиксаторы, живущие в клубеньках корней бобовых растений (клубеньковые бактерии), относящиеся к роду Rhizobium. Azotobacte r. Для того чтобы эти микроорганизмы осуществляли процесс фиксации азота, необходимо присутствие молибдена, железа и кальция. Конечным продуктом фиксации азота является аммиак. В процессе восстановления азота до аммиака участвует мультиферментный комплекс — нитрогеназа. Источником протонов и электронов для восстановления азота служит дыхательная электрон-транспортная цепь. Это указывает на связь усвоения азота атмосферы с процессами дыхания, а также фотосинтеза (источника углеводов). Для восстановления N2 до NH3 требуется шесть электронов, согласно уравнению:N2 + 6е + 2Н+ -> 2NH3 Процесс требует АТФ как источника энергии: затрачивается 25—35 молекул АТФ, т.к нитрогеназа — фермент с низкой субстратной специфичностью. Особенность нитрогеназы заключается и в том, что для работы фермента требуются анаэробные условия. Вместе с тем в клетках высшего растения кислород необходим для поддержания дыхания. Роль леггемоглобина заключается в связывании 02 в организме бактерий и создании условий для работы нитрогеназы. Для образования леггемоглобина необходимы Fe, Сu и Со. Для нормального протекания процесса азотофиксации необходимы Мо и Fe, входящие в состав нитрогеназы. Молибден поддерживает конформацию нитрогеназы, участвует в связывании азота и переносе электронов, а также индуцирует синтез нитрогеназы. Кобальт входит в состав витамина В12, который вовлекается в процесс биосинтеза леггемоглобина. В надземные органы растения-хозяина азотистые вещества передвигаются главным образом в виде амидов (аспарагина, глутамина).
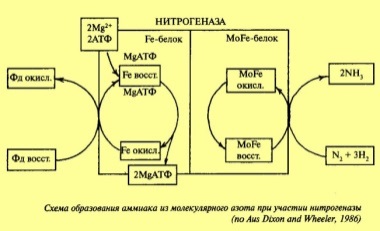
5. Азотный обмен высших растений: восстановление нитратов и пути их усвоения аммиака.
Высшие растения поглощают соединения азота из почвы. Основным источником азотного питания для растений являются нитраты и аммиак. Корневые системы растений хорошо усваивают нитраты, которые, поступая в корни растения, подвергаются ферментативному восстановлению до нитритов и далее до аммиака. Этот процесс происходит главным образом в корнях, однако этой способностью обладают и клетки листьев. Восстановление нитратов до аммиака идет через ряд этапов. На первом этапе нитраты восстанавливаются до нитритов при участии фермента нитратредуктазы:
N03- + 2е -> N02
Нитратредуктаза — это фермент, содержащий в своем составе ФАД, гем и молибден. Фермент локализован в цитозоле, где и протекает процесс восстановления нитратов до нитритов. Поставщиком НАДН являются процесс дыхания и отчасти световые реакции фотосинтеза. Именно поэтому восстановление нитратов тесно связано с дыхательным газообменом и фотосинтезом. Для восстановления нитратов могут быть использованы продукты, образующиеся в процессе нециклического фотофосфорилирования (НАДФН, АТФ). При низкой освещенности, дефиците Fe и Мо активность фермента снижается, и нитраты накапливаются в клетке.
Второй этап — восстановление нитратов до аммиака катализируется ферментом нитритредуктазой:
N02- + 6е -> NH4+
Нитритредуктаза — это фермент, содержащий в качестве простетической группы гем. Активность этого фермента выше, чем нитратредуктазы. Нитритредуктаза локализована в хлоропластах листьев или пропластидах корней. Донором электронов в листьях служит восстановленный ферредоксин, который образуется при функционировании на свету ФС I. Нитриты могут поступать в растение из почвы и подвергаются восстановлению до NH4. Передвижение нитритов из цитоплазмы в хлоропласты стимулируется Са. Осуществляется и обратный процесс — окисление аммонийной формы азота в нитратную, что опровергает широко распространенное мнение об исключительно экзогенном происхождении нитратов в растениях.
Важнейшим источником азотного питания является аммонийный азот. Накопление аммиака в клетках, приводит к нежелательным последствиям. Однако растения обладают способностью обезвреживать аммиак путем присоединения его к органическим кислотам с образованием амидов (глутамина, аспарагина). Растения можно разделить на амидные, образующие амиды — аспарагин и глутамин, и аммиачные, образующие соли аммония. В процессе дыхания в качестве промежуточных продуктов образуются органические кислоты, в том числе а-кетоглутаровая и щавелевоуксусная. Эти кислоты в результате реакции прямого восстановительного аминирования присоединяют аммиак: 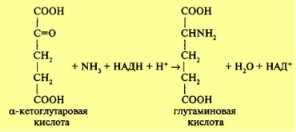 Реакция идет в две стадии. На промежуточном этапе образуется иминокислота. Катализируется реакция ферментом глутаматдегидрогеназой с активной группой НАД. Этот фермент локализован главным образом в митохондриях, но может содержаться в цитозоле и хлоропластах. Аспарагиновая кислота образуется по аналогии с глутаминовой кислотой путем восстановительного аминирования щавелевоуксусной кислоты при участии фермента аспартатдегидрогеназы. На активность глутаминсинтетазы влияет присутствие катионов: Mg2+, Мп2+, Со2+, Са2+. Фермент обнаружен в цитозоле, но может находится и в хлоропластах. Образование аспарагина происходит аналогичным путем. В более молодых органах (листьях) и даже в более молодых клетках образование амидов идет интенсивнее. Роль амидов в растении разнообразна. Это не только форма обезвреживания аммиака, это и транспортная форма азотистых соединений, обеспечивающая отток их из одного органа в другие. Важно, что амиды и их непосредственные предшественники — глутаминовая и аспарагиновая кислоты — являются материалом для построения многих других аминокислот в процессах переаминирования, а также перестройки их углеродного скелета.
Реакция идет в две стадии. На промежуточном этапе образуется иминокислота. Катализируется реакция ферментом глутаматдегидрогеназой с активной группой НАД. Этот фермент локализован главным образом в митохондриях, но может содержаться в цитозоле и хлоропластах. Аспарагиновая кислота образуется по аналогии с глутаминовой кислотой путем восстановительного аминирования щавелевоуксусной кислоты при участии фермента аспартатдегидрогеназы. На активность глутаминсинтетазы влияет присутствие катионов: Mg2+, Мп2+, Со2+, Са2+. Фермент обнаружен в цитозоле, но может находится и в хлоропластах. Образование аспарагина происходит аналогичным путем. В более молодых органах (листьях) и даже в более молодых клетках образование амидов идет интенсивнее. Роль амидов в растении разнообразна. Это не только форма обезвреживания аммиака, это и транспортная форма азотистых соединений, обеспечивающая отток их из одного органа в другие. Важно, что амиды и их непосредственные предшественники — глутаминовая и аспарагиновая кислоты — являются материалом для построения многих других аминокислот в процессах переаминирования, а также перестройки их углеродного скелета.
6. Микроэлементы в жизни растений.
Кроме основных элементов, для роста растений необходим целый ряд так называемых микроэлементов. Микроэлементы усваиваются только при низких концентрациях соответствующих солей. При увеличении дозы они становятся уже ядовитыми для растения. Роль их в жизни растений, подобно витаминам, связана с деятельностью ферментов.
Бор. Для того чтобы растение нормально развивалось, его необходимо снабжать бором. При отсутствии бора приостанавливается рост корней и наземной части. Точки роста отмирают, так как клетки молодой растущей ткани —меристемы перестают делиться. Бор принимает участие в процессе прорастания пыльцы и росте завязи, поэтому при недостатке его резко снижается семенная продукция растений.Бор играет большую роль в передвижении Сахаров; ряд борорганических соединений является активаторами роста.
Медь. Значительная доля меди сосредоточена к хлоропластах. По-видимому, медь катализирует какие-то реакции в фотосинтезе, При недостатке меди хлоропласту оказываются недолговечными. Медь входит в состав ряда окислительных ферментов (поли-фенолксидаза, тирозиназа и др.)Медь играет большую роль в белковом обмене.
Цинк. Цинк входит в состав важного фермента —карбоан-гидразы. Кроме того, цинк участвует в синтезе аминокислоты триптофана, являющегося предшественником ауксинов в растении.
Марганец. Катализирует реакции арбоксилирования и играет важную роль в фотосинтезе и дыхании. Преимущественно он скапливается в листьях и в точках роста. Его наличие способствует окислительным превращениям.Присутствие марганца в питательном растворе повышает дыхание корней, при этом заметно увеличивается усвоение нитратного азота. Особенно характерным свойством марганца является его способность окислять соединения железа.
Молибден. Молибден необходим растениям в чрезвычайно малых количествах. Он катализирует процессы восстановления нитратов и синтеза белковых веществ.
7. Механизм поступления ионов в клетку. Роль клеточной оболочки. Транспорт ионов через мембраны: пассивный и активный.
Поглощение ионов клеткой начинается с их взаимодействия с клеточной стенкой. Ионы могут частично локализоваться в межмицеллярных и межфибриллярных промежутках клеточной стенки, фиксироваться в клеточной стенке электрическими зарядами. Объем клетки, доступный для свободной диффузии ионов, получил название свободного пространства. Свободное пространство включает межклетники, клеточные стенки и промежутки, которые могут возникать между клеточной стенкой и плазмалеммой. Иногда его называют кажущееся свободное пространство (КСП). Термин “кажущееся” означает, что объем зависит от объекта и природы растворенного вещества. Свободное пространство растения получило название апопласт, симпласт — совокупность протопластов всех клеток. Поглощение и выделение веществ в КСП — физико-химический пассивный процесс, не зависимый от температуры и ингибиторов энергетического и белкового обменов. Клеточная стенка обладает свойствами ионообменника, так как в ней адсорбированы ионы Н + и НCO — 3, обменивающиеся в эквивалентных количествах на ионы внешнего раствора. Для того чтобы проникнуть в цитоплазму и включиться в обмен веществ, ионы должны пройти через плазмалемму. Транспорт ионов через мембрану может быть пассивным и активным. Пассивное поглощение не требует затрат энергии и осуществляется путем диффузии по градиенту концентрации вещества, для которого плазмалемма проницаема. Пассивное передвижение ионов определяется не только химическим потенциалом m, как это имеет место при диффузии незаряженных частиц, но и электрическим потенциалом e. Оба потенциала объединяют в виде электрохимического потенциала ` m: ` m = m + nF e, где m — химический, e — электрический, ` m — электрохимический потенциалы, n — валентность иона, F — константа Фарадея. Нужно это?
Электрический потенциал на мембране — трансмембранный потенциал может возникнуть по следующим причинам: 1) если поступление ионов идет по градиенту концентрации, но благодаря разной проницаемости мембраны с большей скоростью поступают катионы, чем анионы. В силу этого на мембране возникает разность электрических потенциалов, что приводит к диффузии противоположно заряженного иона; 2) при наличии на внутренней стороне мембраны белков, фиксирующих определенные ионы; 3) в результате активного транспорта либо катиона, либо аниона, в этом случае противоположно заряженный ион может передвигаться пассивно по градиенту электрического потенциала.
Активный транспорт — это транспорт, идущий против электрохимического градиента с затратой энергии, выделяющейся в процессе метаболизма. С температурой возрастает, яды ингибируют. А увеличение АТФ усиливает процесс поглощения. Активный транспорт ионов через мембрану осуществляется с помощью переносчиков. Ион реагирует со своим переносчиком на поверхности плазмалеммы. Комплекс переносчика с ионом подвижен в самой мембране и передвигается к ее внутренней стороне. Здесь комплекс распадается и ион освобождается во внутреннюю среду, а переносчик передвигается к внешней стороне мембраны. Транспорт с участием переносчиков может идти по градиенту электрохимического потенциала. Это пассивный транспорт, но благодаря переносчикам он идет с большей скоростью, чем обычная диффузия и этот процесс носит название облегченной диффузии. Для использования энергии АТФ должна быть гидролизована: АТФ + НОН ® АДФ + Фн. Катализируется АТФазой. Транспортные АТФазы являются высокомолекулярными липопротеидами. Для растений большое значение имеет Н + — АТФаза (водородный насос или водородная помпа. Перенос ионов водорода сопровождается переносом катионов в обратном направлении. Такой процесс называется антипорт. Вместе с протоном могут двигаться анионы — симпорт. Освобождаемая при распаде АТФ энергия используется для изменения конфигурации самой АТФазы, благодаря чему участок фермента, связывающий определенный ион, поворачивается и оказывается по другую сторону мембраны.
Внутриклеточный транспорт ионов осуществляется благодаря движению цитоплазмы и по каналам эндоплазматического ретикулума. Ионы попадают в вакуоль, если цитоплазма и органеллы уже насыщены ими. Для того, чтобы попасть в вакуоль, ионы должны преодолеть еще один барьер — тонопласт. Транспорт ионов через тонопласт совершается также с помощью переносчиков и требует затраты энергии.
8. Корень как орган поглощения минеральных элементов. Метаболизм корней.
Основной зоной поглощения питательных веществ, снабжающей и надземные органы растения, является зона растяжения клеток и зона корневых волосков. В меристематической зоне нет дифференцированной сосудистой системы. При этом флоэма дифференцируется раньше, и лишь несколько выше по длине корня образуется ксилема. Именно по ксилеме происходит передвижение воды с растворенными питательными веществами. Поэтому основная масса поглощенных меристемой ионов используется в этих же клетках.
Выше зоны корневых волосков расположена зона ветвления корня. В этой зоне поверхность покрыта слоем пробки и в поглощении питательных солей практически не участвует. Различные зоны корня поглощают разные минеральные элементы. Показано, что Са2+ поступает только в апикальные зоны, К+, NH4+, фосфаты абсорбируются всей корневой системой.
Особенности поступления: способность растений к избирательному концентрированию веществ; относительная независимость поступления воды и солей; зависимость от дыхания и фотосинтеза; ускорение процесса под влиянием температуры и света.
В корневой системе различают два объема — апопласт и симпласт. Большая часть коллоидов почвы заряжена отрицательно, на их поверхности в адсорбированном (поглощенном) состоянии находятся катионы. Катионы и анионы, находящиеся в поглощенном состоянии на частицах почвенного поглощающего комплекса, могут обмениваться на ионы, адсорбированные на поверхности клеток корня. Так может осуществляться поступление катионов К+, Са2+, Na+ в обмен на протоны, а также анионов N03-, Р043- и других в обмен на НС03- или анионы органических кислот. Поглощенные ионы адсорбируются на поверхности клеточных оболочек ризодермы. Из адсорбированного состояния ионы могут по коре корня передвигаться двумя путями: по апопласту и симпласту. При поступлении в симпласт ионы проникают через мембрану и далее передвигаются по плазмодесмам к сосудам ксилемы.
Ток воды с растворенными веществами, движущийся по свободному пространству (апопласту), как бы омывает все клетки коры. На всем этом пути могут наблюдаться адсорбция веществ клеточными стенками, поступление ионов в клетки через соответствующие мембраны и включение их в обмен веществ, т. е. метаболизация. Необходимо учесть, что на пути движения по апопласту имеется преграда в виде клеток эндодермы с поясками Каспари. Передвижение через клетки эндодермы возможно, по-видимому, только через цитоплазму. В связи с этим перенос ионов через мембраны клеток эндодермы необходим и также осуществляется с помощью переносчиков. В целом между апопластом и симпластом в корневой системе происходит непрерывное взаимодействие и обмен питательными солями.
9. Экология минерального питания: влияние внешних и внутренних факторов. Физиологические основы применения удобрений.
Влияние внешних условий
При температуре, близкой к 0°С, поглощение солей идет медленно, затем, в пределах до 40°С, оно усиливается. Увеличение температуры на 10°С может вызвать возрастание поглощения в два и даже в три раза. В темноте поглощение солей замедляется и постепенно прекращается, а под влиянием освещения ускоряется. Снижение концентрации кислорода ниже 3% вызывает падение поглощения примерно в два раза. В присутствии легко поглощаемого аниона катионы той же соли поступают быстрее. Ионы с одинаковым зарядом обычно конкурируют между собой.
Влияние внутренних факторов
Зависит от интенсивности дыхания. Процесс дыхания может оказывать влияние на поступление солей в нескольких направлениях. Так, в процессе дыхания выделяющийся углекислый газ в водной среде диссоциирует на ионы Н+ и НС03-. Адсорбируясь на поверхности корня, эти ионы служат обменным фондом для поступающих катионов и анионов. В процессе переноса ионов через мембрану участвуют специфические белки-переносчики, синтез которых находится в зависимости от интенсивности дыхательного процесса.
Рациональное внесение питательных веществ в виде удобрений — мощный фактор повышения урожайности растений. Однако необходимо учитывать, что завышенные дозы удобрений представляют не только бесполезную их трату, но могут привести к ряду весьма вредных последствий. Прежде всего, это может создать повышенную концентрацию почвенного раствора. Большинство культурных растений чувствительны к этому показателю. Повышение содержания какой-либо питательной соли может оказать непосредственное токсическое действие на растительный организм. Для установления обоснованных норм удобрений необходимо учитывать наличие питательных веществ в почве, потребности данного растения, свойства вносимых удобрений, состав корневых выделений, транспирационные коэффициенты, фотосинтез, кислотность почв.

