 2015-02-14
2015-02-14 13199
13199Первичная структура - последовательность расположения мононуклеотидов в полинуклеотидной цепи ДНК и РНК. Нуклеотиды в молекулах нуклеиновых кислот соединены сложноэфирной связью, образованной остатком фосфорной кислоты одного мононуклеотида и 3’-гидроксильной группой остатка пентозы другого мононуклеотида (3’,5’-фосфодиэфирная связь) (рис. 11).
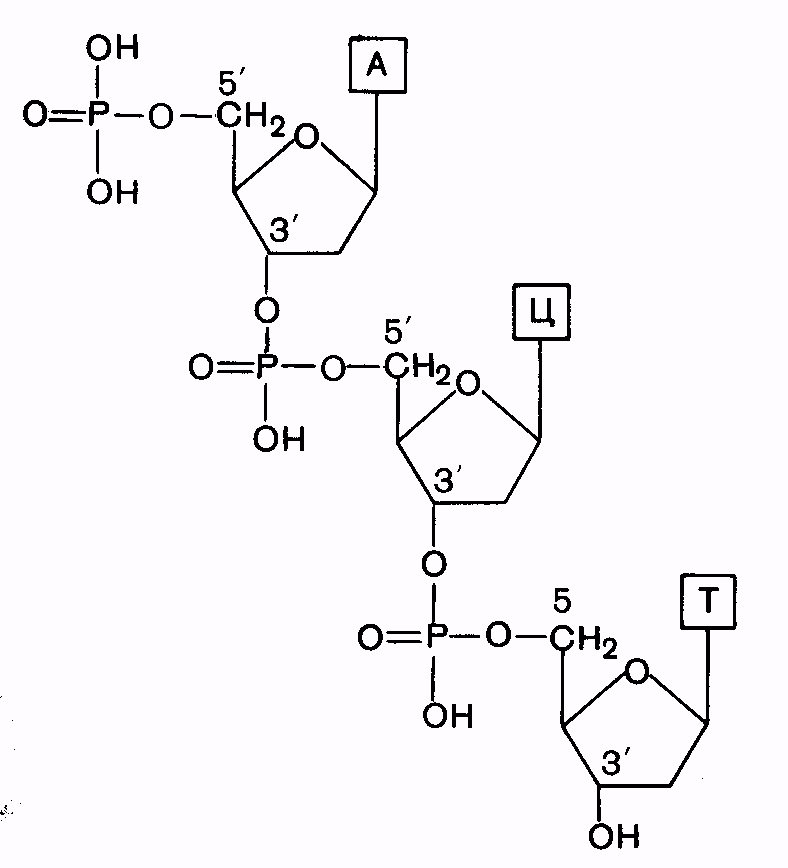 | Рис. 11. Схема первичной структуры ДНК |
Для изучения химического состава НК используют секвенирование – расщепление НК на фрагменты ферментами либо химическими реагентами и анализ продуктов при помощи электрофореза, хроматографии и др.
Вторичная структура
ДНК состоит из двух цепей, закрученных вокруг одной и той же оси, образующих правовращающую спираль. Внутри спирали находятся азотистые основания, снаружи - углеводные компоненты (рис. 12). Обе цепи в молекуле ДНК имеют противоположную полярность: межнуклеотидная связь в одной цепи имеет направление 5’®3’, в другой – 3’®5’.
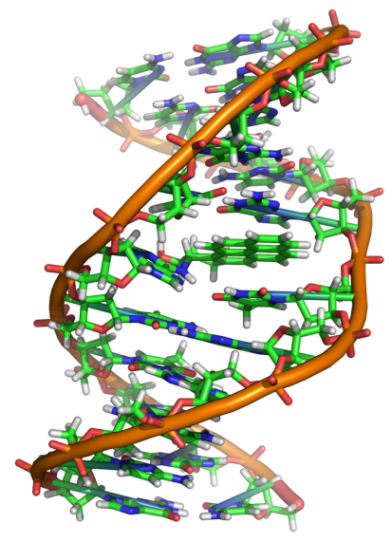 |
| Рис. 12. Вторичная структура ДНК |
В молекуле ДНК основания уложены парами: пуриновые из одной цепи и пиримидиновые из другой. Взаимодействие пар А-Т и Г-Ц называют комплементарностью, а соответствующие азотистые основания – комплементарными (рис. 13). Цепи ДНК комплементарны друг другу. Стабильность А-Т пар обеспечивается двумя водородными связями, а пар Г-Ц-тремя.
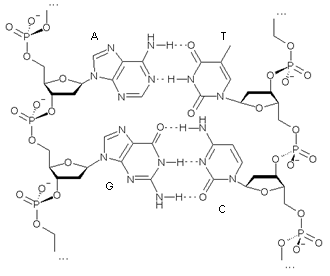 |
| Рис. 13. Комплементарность цепей в ДНК |
Между азотистыми основаниями, собранными в стопку вдоль молекулы ДНК, возникают силы гидрофобных взаимодействий (стэкинг-взаимодействия), которые вносят большой вклад в стабилизацию двойной спирали.
Конфигурация двойной спирали ДНК меняется от количественного содержания воды и ионной силы раствора. Существует более 10 форм ДНК. Методами рентгеноструктурного анализа изучено шесть: А-, В-, С-, D-, Е- и Z-формы.
У А-формы пары комплементарных оснований смещены от оси молекулы к периферии, а в В-форме - близки к оси спирали. В А-форме ДНК является матрицей в транскрипции (синтез РНК на молекуле ДНК), в В-форме - в репликации (синтез ДНК на молекуле ДНК). Параллельно фосфодиэфирному остову в структуре А- и В-форм ДНК имеются большая и малая бороздки – сайты присоединения белков.
Z-форма (зигзагообразная) имеет левозакрученную конфигурацию, в которой фосфатные группы располагаются зигзагообразно вдоль оси молекулы.
SBS-форма ДНК характеризуется отсутствием взаимозакручивания цепей в биспиральную молекулу, они располагаются бок о бок (side by side). Такая форма ДНК обеспечивает легкое распаривание и расхождение цепей, что очень важно при биосинтезе ДНК.
Молекулы РНК построены из одной полинуклеотидной цепи. В этой цепи есть комплементарные участки, которые образуют двойные спирали. При этом соединяются водородными связями пары А-У и Г-Ц. Спирализованные участки РНК (шпильки) содержат 20-30 нуклеотидных пар и чередуются с неспирализованными участками.
Для тРНК Р. Холли предложил модель клеверного листа - спирализация полинуклеотидной цепи самой на себя в строго фиксированных зонах. Особенности структуры тРНК имеют прямое отношение к процессу трансляции, поэтому более подробно они рассмотрены в разделе биосинтеза белка.
Вторичная структура рРНК и мРНК характеризуется спирализацией самой на себя.
Во вторичной структуре ДНК и РНК есть нуклеотидные последовательности, называемые палиндромами (перевернутые повторы), которые служат основой для образования структуры шпилек или для формирования на отдельных участках тройных спиралей.
Третичная структура нуклеиновых кислот: двойная спираль ДНК на некоторых участках может подвергаться дальнейшей спирализации с образованием суперспирали или открытой кольцевой формы. Нативные молекулы тРНК обладают третичной структурой, которая отличается от «листа клевера» компактностью за счет складывания различных частей молекулы.
В растянутом виде ДНК человека имеет около двух метров в длину, но в живой клетке упакована в структуру размером в доли миллиметра.
Нуклеиновые кислоты входят в состав нуклеопротеинов.
Существует два типа НП - дезоксирибонуклеопротеины (ДНП, содержат ДНК) и рибонуклеопротеины (РНП, содержат РНК). ДНП преимущественно находятся в ядре, а РНП - в цитоплазме. В состав НП входят гистоновые и негистоновые белки. Представители НП - рибосомы (комплексы рибосомных РНК с белками), ДНП-хроматин (комплекс ДНК с гистонами и негистоновыми белками).
Гистоны - сильно щелочные белки с невысокой молекулярной массой. Содержат большое количество лизина и аргинина. Пять классов гистонов различаются по размерам, аминокислотному составу и величине заряда. Они принимают участие в структурной организации хроматина, нейтрализуя за счет положительных зарядов аминокислотных остатков отрицательно заряженные фосфатные группы ДНК.
Природа негистоновых белков пока выяснена недостаточно. Их изоэлектрическая точка находится в кислой среде.
Для многих белков хроматина характерны особенности структуры, обеспечивающие их связывание с ДНК: «лейциновая застежка-«молния»; мотив «α-спираль – поворот – α-спираль», «цинковый палец».
В составе нуклеопротеинов НК имеет, как правило, более компактную структуру, чем в изолированном виде.

