2015-02-14
2015-02-14 10451
10451Как уже отмечалось распад олигосахаридов и полисахаридов осуществляется путём гидролиза и фосфоролиза. В первом случае образуются свободные монозы, а во втором – их фосфорные эфиры. Дальнейший обмен моносахаридов идёт такими путями, что используются только фосфорные эфиры моносахаридов. Свободные же монозы фосфорилируются, превращаясь в соответствующие фосфорные эфиры. Реакция фосфорилирования моносахаридов рассматривается в биохимии как реакция их активирования, поскольку фосфорные эфиры моноз гораздо реакционноспособнее, чем свободные моносахариды.
Фосфорилирование моносахаридов осуществляется при взаимодействии их с АТФ и ускоряется специфическими фосфотрансферазами, которые называются киназами. Например, фосфорилирование глюкозы ускоряется ферментом глюкокиназой, полное номенклатурное название которой АТФ: D-глюкоза-6-фосфотрансфераза:
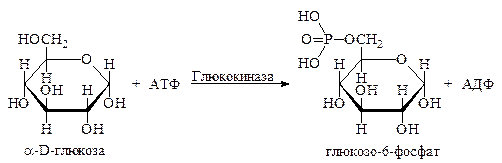
В аналогичную реакцию вступают и другие моносахариды, такие как фруктоза и рибоза. Фруктоза при фосфорилировании образует фруктозо-6-фосфат, а рибоза – рибозо-5-фосфат.
В природе обнаружено более двух десятков индивидуальных фосфотрансфераз, переносящих остатки фосфорной кислоты на те или иные моносахариды. Следовательно, практически любой моносахарид может быть переведён в фосфорный эфир.
Важной особенностью фосфорных эфиров моносахаридов является их способность к изомеризации под действием ферментов изомераз. Причём изомеризоваться они могут двумя путями: путём внутримолекулярного переноса атомов водорода (взаимопревращения альдоз и кетоз) и путём изменения пространственного расположения атомов и атомных групп в молекуле (стереоизомеризация). Примерами таких реакций служат переход глюкозо-6-фосфата во фруктозо-6-фосфат и превращения рибозо-5-фосфата.
Изомеризация глюкозо-6-фосфата во фруктозо-6-фосфат интенсивно осуществляется в мышечной ткани и приводит к установлению равновесия между этими двумя эфирами в соотношении 2: 1:
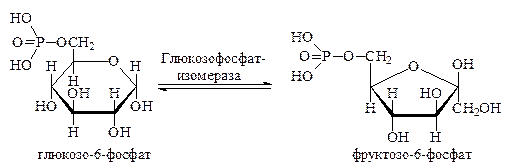
Рибозо-5-фосфат вначале превращается в свою кето-форму – рибулозо-5-фосфат, а рибулозо-5-фосфат, подвергаясь эпимеризации – в ксилулозо-5-фосфат. Превращения катализируются соответствующими ферментами:
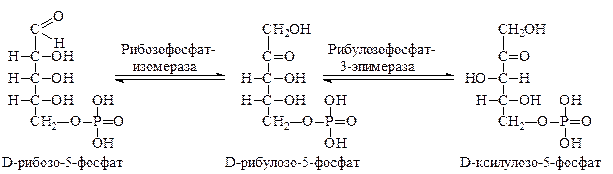
Очень легко процессы перестройки одних моносахаридов в другие протекают, если фосфорные эфиры моносахаридов соединены с уридин-5¢-фосфатом. Пример реакции такого рода – обратимое превращение УДФ-глюкозы в УДФ-галактозу:
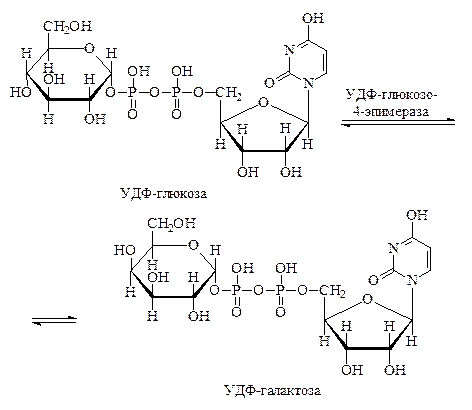
Как итог рассмотренным процессам, можно сказать, что в живом организме любой моносахарид и его фосфорный эфир может превратиться в любой другой, изомерный ему. Это играет огромную роль, поскольку процесс обмена моносахаридов протекает только через некоторые их ключевые формы: глюкозо-6-фосфат и рибулозо-5-фосфат.
Поэтому следующий вопрос, который необходимо разобрать для понимания метаболизма моносахаридов, – обмен глюкозо-6-фосфата.
Обмен глюкозо-6-фосфата
Глюкозо-6-фосфат образуется в организме разными путями.
Во-первых, он может синтезироваться путём фосфорилирования глюкозы за счёт её взаимодействия с АТФ.
Во-вторых, он образуется в результате изомеризации фосфорных эфиров других гексоз.
В-третьих, он получается из глюкозо-1-фосфата – продукта фосфоролиза олиго- и полисахаридов.
Первые две реакции мы уже рассматривали ранее. Что касается преобразования глбкозо-1-фосфата в глюкозо-6-фосфат, то эта реакция протекает в два этапа и ускоряется ферментом фосфоглюкомутазой. Активный центр фосфоглюкомутазы содержит остаток фосфосерина, с которого остаток фосфорной кислоты передается на глюкозо-1-фосфат с образованием глюкозо-1,6-дифосфата и дефосфорилированного фермента. Затем дефосфорилированная фосфоглюкомутаза, взаимодействуя с глюкозо-1,6-дифосфатом, снова превращается в фосфопротеин, однако остаток фосфорной кислоты она отщепляет не от 6-го углеродного атома фосфорного эфира, а от первого. В результате этой реакции фермент возвращается в прежнее состояние и высвобождается глюкозо-6-фосфат:
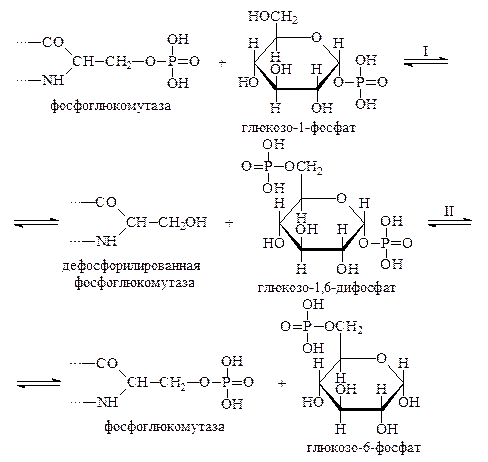
Описанная реакция обратима, однако её равновесие сдвинуто в сторону образования глюкозо-6-фосфата. Поэтому содержание глюкозо-1-фосфата в тканях организмов обычно не превышает 3–4% от общего количества гексозомонофосфатов.
Далее глюкозо-6-фосфат подвергается в организме различным превращениям. Некоторая доля его распадается до углекислого газа и воды. При этом многократно повторяются реакции окисления как самого глюкозо-6-фосфата, так и продуктов его дальнейшего распада. Сопряжённо с его окислением осуществляется синтез АТФ из АДФ и H3PO4. Таким образом глюкозо-6-фосфат является, во-первых, источником энергии для организма.
Вместе с тем значительная часть промежуточных продуктов распада глюкозо-6-фосфата используется для синтеза аминокислот, нуклеотидов, глицерина и ВЖК, стеролов и т.д. В частности, пировиноградная кислота используется для синтеза аланина; 3-фосфоглицериновая кислота и фосфоенолпировиноградная кислота идут на синтез фенилаланина, тирозина, триптофана и серина; пировиноградная кислота, включаясь в цикл Кребса, превращается в щавелевоуксусную и a-кетоглутаровую, которые дают начало аспарагиновой и глутаминовой кислотам; рибозо-5-фосфат служит для синтеза гистидина, пиримидиновых и пуриновых нуклеотидов:
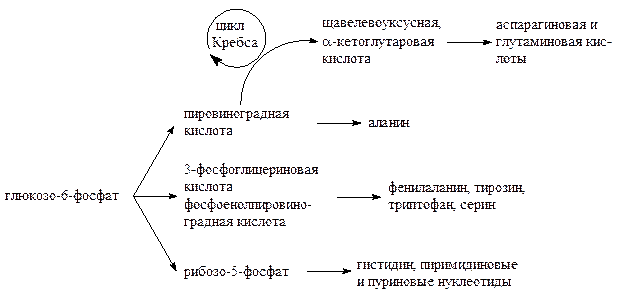
Таким образом, второе предназначение обмена глюкозо-6-фосфата – снабжать организм строительным материалом для синтеза новых органических соединений, используемых в процессе жизнедеятельности.
Существует три пути распада глюкозо-6-фосфата: дихотомический, апотомический и путь Этнера-Дудорова. Рассмотрим каждый из них.
Дихотомический путь распада глюкозо-6-фосфата. Называется так потому, что на определённой стадии его происходит распад шестиуглеродной молекулы на две трёхуглеродные.
Вступая на дихотомический путь распада, глюкозо-6-фосфат прежде всего изомеризуется и превращается во фруктозо-6-фосфат. Фруктозо-6-фосфат далее фосфорилируется по 1-му углеродному атому, образуя фруктозо-1,6-дифосфат. Первую реакцию мы уже рассмотрели чуть раньше. Вторая же реакция ускоряется ферментом фосфофруктокиназой, практически необратима, очень медленна (в 500 раз медленнее, чем реакция, катализируемая глюкозофосфат-изомеразой) и лимитирует весь процесс дихотомического распада:
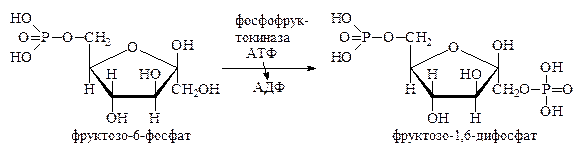
Фруктозо-1,6-дифосфат подвергается далее дихотомическому распаду на две фосфотриозы – фосфодиоксиацетон и 3-фосфоглицериновый альдегид:
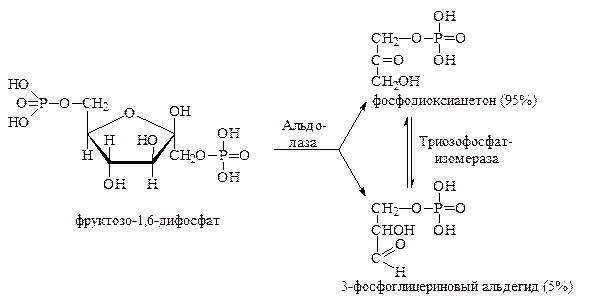
Фосфодиоксиацетон и 3-фосфоглицериновый альдегид изомерны друг другу и могут превращаться один в другой при помощи фермента триозофосфатизомеразы. Хотя первоначально масса фосфодиоксиацетона составляет 95% от массы образующихся фосфотриоз, в дальнейших превращениях участвует только 3-фосфоглицериновый альдегид. По мере расходования 3-фосфоглицеринового альдегида, его убыль пополняется за счёт фосфодиоксиацетона, который практически нацело в него превращается. В конечном счёте из каждой молекулы фруктозо-1,6-дифосфата фактически возникают две молекулы 3-фосфоглицеринового альдегида, который претерпевает дальнейший многостадийный распад:
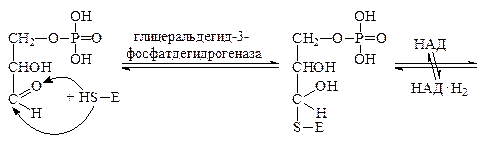
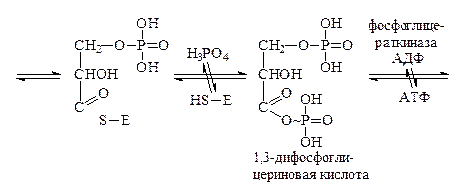
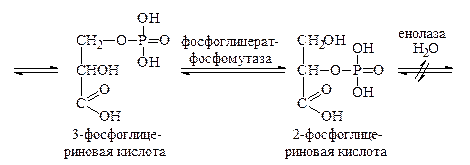
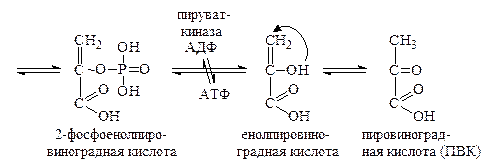
Сперва 3-фосфоглицериновый альдегид окисляется с помощью фермента глицеральдегид-3-фосфатдегидрогеназы. Активный центр каждой субъединицы этого фермента содержит 4 радикала цистеина со свободными HS-группами. Сначала каждая молекула фосфоглицеринового альдегида присоединяется к ферменту по одной из тиольных групп, затем в действие вступает кофермент НАД+, входящий в состав фермента глицеральдегид-3-фосфатдегидрогеназы. Он отнимает от молекулы 3-фосфоглицеринового альдегида два атома водорода, окисляя его. В этот момент связь между образовавшимся остатком фосфоглицериновой кислоты и ферментом становится макроэргической. Эта связь спонтанно распадается в присутствии H3PO4 с образованием 1,3-дифосфоглицериновой кислоты. Фермент приэтом отщепляется.
Затем происходит субстратное фосфорилирование, в результате которого остаток фосфорной кислоты из 1,3-дифосфоглицериновой кислоты переносится на АДФ с образованием АТФ. Процесс катализируется ферментом фосфоглицераткиназой. Далее образовавшаяся 3-фосфоглицериновая кислота изомеризуется под действием фосфоглицерат-фосфомутазы в 2-фосфоглицериновую кислоту, которая, теряя воду, превращается в 2-фосфоенолпировиноградную кислоту. Связь с остатком фосфорной кислоты становится при этом макроэргической. Фосфоенолпировиноградная кислота вступает в реакцию субстратного фосфорилирования с АДФ с образованием АТФ и енолпировиноградной кислоты. Последняя изомеризуется в процессе кето-енольной таутомерии в более стабильную пировиноградную кислоту.
Таким образом, конечным продуктом распада глюкозо-6-фосфата по дихотомическому пути является пировиноградная кислота. Её дальнейшие превращения мы рассмотрим позднее. Теперь перейдём к апотомическому пути распада глюкозо-6-фосфата.
Апотомический путь распада глюкозо-6-фосфата. В отличие от дихотомического пути, он начинается не с реакции фосфорилирования, а с реакции окисления. Глюкозо-6-фосфат под действием фермента глюкозо-6-фосфатдегидрогеназы превращается в 6-фосфоглюконолактон. Акцептором водорода служит НАДФ+, которая является коферментом глюкозо-6-фосфатдегидрогеназы:
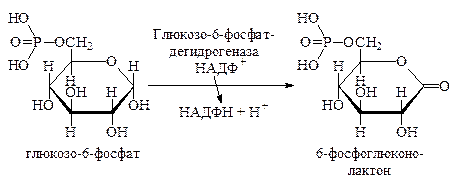
Образовавшийся лактон далее гидролизуется до 6-фосфоглюконовой кислоты при участии фермента глюконолактоназы. 6-Фосфоглюконовая кислота далее претерпевает окислительное декарбоксилирование, превращаясь в рибулозо-5-фосфат:
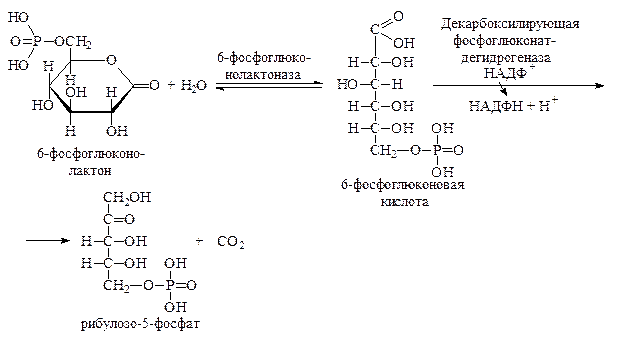
Рибулозо-5-фосфат является одним из центральных веществ в углеводном обмене. Его дальнейшие превращения протекают достаточно сложно. Он многократно изомеризуется, переходя в рибозо-5-фосфат и ксилулозо-5-фосфат; вступает в трансальдолазные и транскетолазные реакции, заключающиеся в переносе двух- и трёхуглеродных фрагментов от одного фосфорного эфира углевода к другому. В конечном счёте из него снова образуется глюкозо-6-фосфат. При этом из каждых 6 молекул рибулозо-5-фосфата получается 5 молекул глюкозо-6-фосфата. Таким образом, суммарный эффект всех реакций апотомического распада глюкозо-6-фосфата сводится к тому, что из каждых 6 его молекул распадается одна. Это можно выразить следующими уравнениями:
6 мол-л глюкозо-6-фосфата + 12НАДФ+ + 7H2O ® 6CO2 + 12НАДФН + 12Н+ + H3PO4 + 5 мол-л глюкозо-6-фосфата
глюкозо-6-фосфат + 12НАДФ+ + 7H2O ® 6CO2 + 12НАДФН + 12Н+ + H3PO4
Поскольку огромную роль в апотомическом распаде играют превращения фосфорных эфиров пентоз, то такой путь обмена углеводов называется по-другому пентозофосфатным циклом. Эволюционно он относится к числу наиболее древних. Однако в процессе эволюции дихотомический путь распада стал преобладающим и сейчас пентозофосфатный цикл у высших животных занимает скромное место.
Путь Этнера-Дудорова. Этот третий путь обмена глюкозо-6-фосфата, названный по имени ученых, открывших его в 1952 г., характерен в основном для микроорганизмов, у которых при его посредстве распадается до 50% глюкозы.
Первые стадии этого пути до образования 6-фосфоглюконовой кислоты включительно, полностью повторяют апотомический путь. Затем 6-фосфоглюконовая кислота окисляется без декарбоксилирования в 2-кето-6-фосфоглюконовую кислоту, которая, восстанавливаясь по 3-му углеродному атому, переходит в 2-кето-3-дезокси-6-фосфоглюконовую кислоту, расщепляющуюся до пировиноградной кислоты и 3-фосфоглицеринового альдегида:
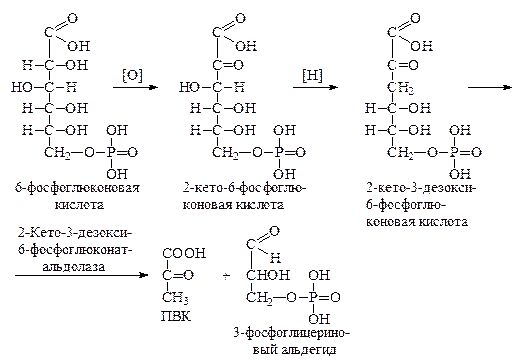
Оба конечных продукта распада попадают в общий метаболический фонд и подвергаются дальнейшим превращениям.
Пировиноградная кислота, образующаяся в этом и дихотомическом путях обмена глюкозо-6-фосфата, является важнейшим метаболитом организмов и претерпевает дальнейшие превращения, которые мы и рассмотрим далее.