форменные элементы крови человека в мазке. 1 – эритроцит, 2 – сегментоядерный нейтрофильный гранулоцит, 3 – палочкоядерный нейтрофильный гранулоцит, 4 – юный нейтрофильный гранулоцит, 5 – эозинофильный гранулоцит, 6 – базофильный гранулоцит, 7 – большой лимфоцит, 8 – средний лимфоцит, 9 – малый лимфоцит, 10 – моноцит, 11 – тромбоциты (кровяные пластинки). Электронная микрофотография гемолиза эритроцитов и образование их “теней” 1 – дискоцит, 2 – эхиноцит, 3 – “тени” (оболочки) эритроцитов.
К форменным элементам крови относятся эритроциты, лейкоциты и тромбоциты. Эритроциты выполняют в организме следующие функции: 1) основной функцией является дыхательная – перенос кислорода от альвеол легких к тканям и углекислого газа от тканей к легким; 2) регуляция рН крови благодаря одной из мощнейших буферных систем крови – гемоглобиновой; 3) питательная – перенос на своей поверхности аминокислот от органов пищеварения к клеткам организма; 4) защитная – адсорбция на своей поверхности токсических веществ; 5) участие в процессе свертывания крови за счет содержания факторов свертывающей и противосвертывающей систем крови; 6) эритроциты являются носителями разнообразных ферментов (холинэстераза, угольная ангидраза, фосфатаза) и витаминов (В1, В2, В6, аскорбиновая кислота); 7) эритроциты несут в себе групповые признаки крови. Эритроциты составляют более 99% клеток крови. Они составляют 45% объема крови. Эритроциты - это красные кровяные тельца, имеющие форму двояковогнутых дисков диаметром от 6 до 9 мкм, а толщиной 1 мкм с увеличением к краям до 2,2 мкм. Эритроциты такой формы называются нормоцитами. Особая форма эритроцитов приводит к увеличению диффузионной поверхности, что способствует лучшему выполнению основной функции эритроцитов – дыхательной. Специфическая форма обеспечивает также прохождение эритроцитов через узкие капилляры. Кровь имеет красный цвет благодаря присутствующему в эритроцитах белку, который называется гемоглобин. Именно гемоглобин связывает кислород и разносит его по всему организму, обеспечивая дыхательную функцию и поддержание рН крови. Гемоглобин - белок, образованный четырьмя цепями аминокислот. Каждая цепь присоединяется к молекулярной группе, группе гема, которая имеет один атом железа, фиксирующий молекулу кислорода. При этом валентность железа, к которому присоединяется кислород, не изменяется, т.е. железо остается двухвалентным. Гемоглобин, присоединивший к себе кислород, превращается в ярко красное вещество оксигемоглобин. Это соединение непрочное. В виде оксигемоглобина переносится большая часть кислорода. После высвобождения кислорода возникает более темное вещество, называемое дезоксигемоглобин. У мужчин в крови содержится в среднем 130 – 1б0 г/л гемоглобина, у женщин – 120 – 150 г/л. В клинических условиях принято вычислять степень насыщения эритроцитов гемоглобином. Это так называемый цветовой показатель. В норме он равен 1. Такие эритроциты называются нормохромными. При цветовом показателе более 1,1 эритроциты гиперхромные, менее 0,85 – гипохромные. Цветовой показатель важен для диагностики анемий различной этиологии. Содержание эритроцитов в крови обозначают их числом в одном кубическом миллиметре. В норме в крови у мужчин содержится 4,0 – 5,0х10"/л, или 4 млн – 5 млн эритроцитов в 1 мкл, у женщин – 4,5х10"/л, или 4,5 млн в 1 мкл. Повышение количества эритроцитов в крови называется эритроцитозом, уменьшение эритропенией. Образование эритроцитов происходит в костном мозге путем эритропоэза. Образование идет непрерывно, потому что каждую секунду макрофаги селезенки уничтожают около двух миллионов отживших эритроцитов, которые нужно заменить. Кровь снабжается клетками в основном при помощи красного костного мозга (тельца миелоидного происхождения). Поэтому у детей практически весь костный мозг-красный, в то время как у взрослого человека его процент составляет только половину, и только в определенных костях производится кровь. Когда лимфоциты переходят в лимфатические узлы, образуются лимфоциты В, участвующие в выработке антител, а когда переходят в тимус, образуются лимфоциты Т, вызывающие отторжения при пересадке органов. Но каково происхождение крови? Несмотря на то что это еще довольно неясный вопрос, в настоящее время считается, что все клетки крови восходят к одной единственной изначальной клетке - материанской полипотентной клетке, которая порождает различные типы клеток и может воспроизводить сама себя. От нее происходят унопотентные материнские клетки, вынужденные дифференцироваться на эритроциты, лейкоциты или кровяные пластинки. Этот процесс происходит примерно на третьей неделе жизни человеческого зародыша. И только к четвертому месяцу начинают проявлять активность костный мозг и лимфатические органы. Для образования эритроцитов требуются железо и ряд витаминов. Железо организм получает из гемоглобина разрушающихся эритроцитов и с пищей. Для образования эритроцитов требуются витамин В12 (цианокобаламин) и фолиевая кислота. Для нормального эритропоэза необходимы микроэлементы - медь, никель, кобальт, селен. Физиологическими регуляторами эритропоэза являются эритропоэтины, образующиеся главным образом в почках, а также в печени, селезенке и в небольших количествах постоянно присутствующие в плазме крови здоровых людей. Эритропоэтины усиливают пролиферацию клеток-предшественников эритроидного ряда – КОЕ-Э (колониеобразующая единица эритроцитарная) и ускоряют синтез гемоглобина. Они стимулируют синтез информационной РНК, необходимой для образования энзимов, которые участвуют в формировании гема и глобина. Эритропоэтины увеличивают также кровоток в сосудах кроветворной ткани и увеличивают выход в кровь ретикулоцитов. Продукция эритропоэтинов стимулируется при гипоксии различного происхождения: пребывание человека в горах, кровопотеря, анемия, заболевания сердца и легких. Эритропоэз активируется мужскими половыми гормонами, что обусловливает большее содержание эритроцитов в крови у мужчин, чем у женщин. Стимуляторами эритропоэза являются соматотропный гормон, тироксин, катехоламины, интерлейкины. Торможение эритропоэза вызывают особые вещества – ингибиторы эритропоэза, образующиеся при увеличении массы циркулирующих эритроцитов, например у спустившихся с гор людей. Тормозят эритропоэз женские половые гормоны (эстрогены), кейлоны. Симпатическая нервная система активирует эритропоэз, парасимпатическая – тормозит. Нервные и эндокринные влияния на эритропоэз осуществляются, по-видимому, через эритропоэтины. Об интенсивности эритропоэза судят по числу ретикулоцитов– предшественников эритроцитов. В норме их количество составляет 1 – 2%. Созревшие эритроциты циркулируют в крови в течение 100 – 120 дней. Разрушение эритроцитов происходит в печени, селезенке, в костном мозге посредством клеток мононуклеарной фагоцитарной системы. Продукты распада эритроцитов также являются стимуляторами кроветворения. Процесс разрушения оболочки эритроцитов и выход гемоглобина в плазму крови называется гемолизом. При этом плазма окрашивается в красный цвет и становится прозрачной – “лаковая кровь”. Различают несколько видов гемолиза. Осмотический гемолиз может возникнуть в гипотонической среде. Концентрация раствора NаСl, при которой начинается гемолиз, носит название осмотической резистентности эритроцитов, Для здоровых людей границы минимальной и максимальной стойкости эритроцитов находятся в пределах от 0,4 до 0,34%. Химический гемолиз может быть вызван хлороформом, эфиром, разрушающими белково-липидную оболочку эритроцитов. Биологический гемолиз встречается при действии ядов змей, насекомых, микроорганизмов, при переливании несовместимой крови под влиянием иммунных гемолизинов. Температурный гемолиз возникает при замораживании и размораживании крови в результате разрушения оболочки эритроцитов кристалликами льда. Механический гемолиз происходит при сильных механических воздействиях на кровь, например встряхивании ампулы с кровью. Скорость оседания эритроцитов (СОЭ) у здоровых мужчин составляет 2 – 10 мм в час, у женщин – 2 – 15 мм в час. СОЭ зависит от многих факторов: количества, объема, формы и величины заряда эритроцитов, их способности к агрегации, белкового состава плазмы. В большей степени СОЭ зависит от свойств плазмы, чем эритроцитов.
белкового происхождения, чужеродных белков, а также комплекса антиген-антитело. Эозинофилы продуцируют фермент гистаминазу, который разрушает гистамин, освобождающийся из поврежденных базофилов и тучных клеток при различных аллергических состояниях, глистных инвазиях, аутоиммунных заболеваниях. Эозинофилы осуществляют противоглистный иммунитет, оказывая на личинку цитотоксическое действие. Поэтому при этих заболеваниях увеличивается количество эозинофилов в крови (эозинофилия). Эозинофилы продуцируют плазминоген, который является предшественником плазмина – главного фактора фибринолитической системы крови. Содержание эозинофилов в периферической крови подвержено суточным колебаниям, что связано с уровнем глюкокортикоидов. В конце второй половины дня и рано утром их на 20~ меньше среднесуточного уровня, а в полночь – на 30% больше. Базофилы продуцируют и содержат биологически активные вещества (гепарин, гистамин и др.), чем и обусловлена их функция в организме. Гепарин препятствует свертыванию крови в очаге воспаления. Гистамин расширяет капилляры, что способствует рассасыванию и заживлению. В базофилах содержатся также гиалуроновая кислота, влияющая на проницаемость сосудистой стенки; фактор активации тромбоцитов (ФАТ); тромбоксаны, способствующие агрегации тромбоцитов; лейкотриены и простагландины. При аллергических реакциях (крапивница, бронхиальная астма, лекарственная болезнь) под влиянием комплекса антиген-антитело происходит дегрануляция базофилов и выход в кровь биологически активных веществ, в том числе гистамина, что определяет клиническую картину заболеваний. Моноциты обладают выраженной фагоцитарной функцией. Это самые крупные клетки периферической крови и их называют макрофагами. Моноциты находятся в крови 2-3 дня, затем они выходят в окружающие ткани, где, достигнув зрелости, превращаются в тканевые макрофаги (гистиоциты). Моноциты способны фагоцитировать микробы в кислой среде, когда нейтрофилы не активны. Фагоцитируя микробы, погибшие лейкоциты, поврежденные клетки тканей, моноциты очищают место воспаления и подготавливают его для регенерации. Моноциты синтезируют отдельные компоненты системы комплемента. Активированные моноциты и тканевые макрофаги продуцируют цитотоксины, интерлейкин (ИЛ-1), фактор некроза опухолей (ФНО), интерферон, тем самым осуществляя противоопухолевый, противовирусный, противомикробный и противопаразитарный иммунитет; участвуют в регуляции гемопоэза. Макрофаги принимают участие в формировании специфического иммунного ответа организма. Они распознают антиген и переводят его в так называемую иммуногенную форму (презентация антигена). Моноциты продуцируют как факторы, усиливающие свертывание крови (тромбоксаны, тромбопластины), так и факторы, стимулирующие фибринолиз (активаторы плазминогена). Лимфоциты являются центральным звеном иммунной системы организма. Они осуществляют формирование специфического иммунитета, синтез защитных антител, лизис чужеродных клеток, реакцию отторжения трансплантата, обеспечивают иммунную память. Лимфоциты образуются в костном мозге, а дифференцировку проходят в тканях. Лимфоциты, созревание которых происходит в вилочковой железе, называются Т-лимфоцитами (тимусзависимые). Различают несколько форм Т-лимфоцитов. Т–киллеры (убийцы) осуществляют реакции клеточного иммунитета, лизируя чужеродные клетки, возбудителей инфекционных заболеваний, опухолевые клетки, клетки-мутанты. Т-хелперы (помощники), взаимодействуя с В-лимфоцитами, превращают их в плазматические клетки, т.е. помогают течению гуморального иммунитета. Т-супрессоры (угнетатели) блокируют чрезмерные реакции В-лимфоцитов. Имеются также Т-хелперы и Т-супрессоры, регулирующие клеточный иммунитет. Т-клетки памяти хранят информацию о ранее действующих антигенах. В-лимфоциты (бурсозависимые) проходят дифференцировку у человека в лимфоидной ткани кишечника, небных и глоточных миндалин. В-лимфоциты осуществляют реакции гуморального иммунитета. Большинство В-лимфоцитов являются антителопродуцентами. В-лимфоциты в ответ на действие антигенов в результате сложных взаимодействий с Т-лимфоцитами и моноцитами превращаются в плазматические клетки. Плазматические клетки вырабатывают антитела, которые распознают и специфически связывают соответствующие антигены. Различают 5 основных классов антител, или иммуноглобулинов: JgA, Jg G, Jg М, JgD, JgЕ. Среди В-лимфоцитов также выделяют клетки-киллеры, хелперы, супрессоры и клетки иммунологической памяти. О-лимфоциты (нулевые) не проходят дифференцировку и являются как бы резервом Т- и В-лимфоцитов. Лейкоциты образуются в разных органах тела: в костном мозге, селезенке, тимусе, подмышечных лимфатических узлах, миндалинах и пластинках Пэйе, в слизистой оболочке желудка. Процесс образования лейкоцитов, известный как лейкопоэз, может быть различным. С одной стороны, происходит процесс, порождающий гранулоциты: унопотентная материнская клетка претерпевает первое преобразование и превращается в миелобласт, с почти круглым ядром, а затем делится на миелоциты, с собственными признаками, которые приведут соответственно к образованию базофилов, нейтрофилов и эозинофилов. Моноциты всегда сохраняют признаки первичной клетки, поэтому они могут образовываться как при последовательных преобразованиях унопотентной материнской клетки, так и непосредственно из полипотентной материнской клетки. Лейкоциты делятся на две большие группы: гранулоциты и агранулоциты в зависимости от того, наблюдается или нет зернистость в их цитоплазме. У первых имеется ядро различных форм, они осуществляют фагоцитоз. Самые многочисленные и активные - это нейтрофилы (70% от общего числа); кроме них имеются базофилы (1%) и эозинофилы (4%). Незернистые лейкоциты - это моноциты, большего размера и с большой фагоцитарной активностью, и лимфоциты, подразделяющиеся на малые (90%) и большие (остальные 10%).
Тромбоциты, или кровяные пластинки – плоские клетки неправильной округлой формы диаметром 2 – 5 мкм. Тромбоциты человека не имеют ядер - это фрагменты клеток, которые меньше половины эритроцита. Количество тромбоцитов в крови человека составляет 180 – 320х10'/л, или 180 000 – 320 000 в 1 мкл. Имеют место суточные колебания: днем тромбоцитов больше, чем ночью. Увеличение содержания тромбоцитов в периферической крови называется тромбоцитозом, уменьшение – тромбоцитопенией.
Тромбоциты, прилипшие к стенке аорты в зоне повреждения эндотелиального слоя.
Главной функцией тромбоцитов является участие в гемостазе. Тромбоциты помогают "ремонтировать" кровеносные сосуды, прикрепляясь к поврежденным стенкам, а также участвуют в свертывании крови, которое предотвращает кровотечение и выход крови из кровеносного сосуда. Способность тромбоцитов прилипать к чужеродной поверхности (адгезия), а также склеиваться между собой (агрегация) происходит под влиянием разнообразных причин. Тромбоциты продуцируют и выделяют ряд биологически активных веществ: серотонин (вещество, вызывающее сужение кровеносных сосудов уменьшение кровотока), адреналин, норадреналин, а также вещества, получившие название пластинчатых факторов свертывания крови. Так у тромбоцитов есть различные белки, способствующие коагуляции крови. Когда лопается кровеносный сосуд, тромбоциты прикрепляются к стенкам сосуда и частично закрывают брешь, выделяя так называемый тромбоцитарный фактор III, который начинает процесс свертывания крови путем превращения фибриногена в фибрин. Тромбоциты способны выделять из клеточных мембран арахидоновую кислоту и превращать ее в тромбоксаны, которые, в свою очередь, повышают агрегационную активность тромбоцитов. Эти реакции происходят под действием фермента циклооксигеназы.Тромбоциты способны к передвижению за счет образования псевдоподий и фагоцитозу инородных тел, вирусов, иммунных комплексов, тем самым, выполняя защитную функцию. Тромбоциты содержат большое количество серотонина и гистамина, которые влияют на величину просвета и проницаемость капилляров, определяя тем самым состояние гистогематических барьеров. Тромбоциты образуются в красном костном мозге из гигантских клеток мегакариоцитов. Унопотентная клетка претерпевает неполное деление, потому что ядро делится, а цитоплазма нет. В результате образуется мегакариобласт, от цитоплазмы которого в конце отделяются пластинки.Продукция тромбоцитов регулируется тромбоцитопоэтинами. Тромбоцитопоэтины образуются в костном мозге, селезенке, печени. Различают тромбоцитопоэтины кратковременного и длительного действия. Первые усиливают отщепление тромбоцитов от мегакариоцитов и ускоряют их поступление в кровь. Вторые способствуют дифференцировке и созреванию мегакариоцитов.Продолжительность жизни тромбоцитов составляет от 5 до 11 дней. Разрушаются кровяные пластинки в клетках системы макрофагов.Активность тромбоцитопоэтинов регулируется интерлейкинами (ИЛ-6 и ИЛ-11). Количество тромбоцитопоэтинов повышается при воспалении, необратимой агрегации тромбоцитов.
Яичник – парная эндокринная железа овальной формы в среднем длиной 4 см, шириной 2,5 см, толщиной 1,5 см в репродуктивном возрасте, масса её у половозрелой женщины составляет 6-8 г. Яичник покрыт зародышевым эпителием. Он располагается на заднем листке широкой связки матки и прикреплен к ней при помощи небольшой брыжейки, а к матке прикрепляется при помощи собственной связки яичника. В яичнике различают наружный зародышевый, корковый и мозговой слои. Гистологическое строение яичников Наибольшая сложность строения у яичника. Это динамичный орган, в котором происходят постоянные изменения, связанные с гормональным статусом. Развивается из материала полового валика, который закладывается на 4-й неделе эмбриогенеза на медиальной поверхности почек. Он образован целомическим эпителием (из висцерального листка спланхнотома) и мезенхимы. Это индифферентная стадия развития (без половых различий). Специфические различия наступают на 7-8-й неделе. Этому предшествует появление в области полового валика первичных половых клеток – гоноцитов. Они содержат много гликогена в цитоплазме. Из стенки желточного мешка гоноциты по мезенхиме или с током крови попадают в половые валики, встраиваются в эпителиальную пластинку. С этого момента развитие женских и мужских половых желез различается. Образуются яйценосные шары – образования, состоящие из нескольких овогоний, окруженных одним слоем плоских эпителиальных клеток. Затем тяжи мезенхимы разделяют эти шары на более мелкие. Формируются примордиальные фолликулы, состоящие из одной половой клетки, окруженной одним слоем плоских фолликулярных эпителиоцитов. Несколько позже формируются корковое и мозговое вещество. В эмбриональном периоде в яичнике заканчивается период размножения овогенеза и начинается стадия роста, являющаяся самой продолжительной (несколько лет). Овогоний превращается в овоцит первого порядка. Белочная оболочка яичника, соединительнотканная строма, интерстициальные клетки дифференцируются из окружающей мезенхимы. Строение яичника взрослого организма в репродуктивный период. Поверхность яичника покрыта мезотелием, под которым располагается оболочка, образованная плотной соединительной тканью – белочной оболочкой. Под ней лежит корковое вещество, а в центре – мозговое. Мозговое вещество образовано рыхлой соединительной тканью, в которой имеются химосные клетки, продуцирующие гормоны – андрогены. В корковом веществе большое количество кровеносных, лимфатических сосудов и нервных элементов. Основу (строму) коркового вещества образует рыхлая соединительная ткань. В строме в большом количестве располагаются различные фолликулы, желтые и белые тела на разных стадиях развития. В течение репродуктивного периода в яичнике происходит рост овоцита первого порядка в фолликуле. Фолликулы созревают. Последовательные стадии развития фолликула: Самый молодой (их очень много – 30-400 тыс.) – примордиальный фолликул, образованный овоцитом первого порядка, вокруг которого расположен один слой плоских фолликулярных эпителиоцитов, которые выполняют защитную и трофическую функции. Фолликулы располагаются на периферии. На разных стадиях онтогенеза происходит гибель женских половых клеток — атрезия. Первичные фолликулы. Половые клетки несколько больше. На периферии овоцитов первого порядка особая оболочка – блестящая. Вокруг располагается один слой кубических или призматических фолликулярных эпителиоцитов. Прозрачная (блестящая) оболочка образована гликопротеидами. В образовании ее принимают участие овоциты первого порядка. В блестящей оболочке находятся радиально расположенные поры, в которые проникают микроворсинки овоцита и цитоплазматические отростки фолликулярных эпителиоцитов. Вторичные фолликулы. Их образование уже связано с гормональным фоном (воздействие ФСГ). Под его влиянием фолликулярные эпителиоциты начинают усиленно делиться. Вокруг овоцита первого порядка формируется многослойный фолликулярный эпителий. Образование вторичных фолликулов наступает в период полового созревания. Фолликулярный эпителий синтезирует фолликулярную жидкость, которая содержит эстрогены. Образуется полость — пузырчатый фолликул, который постепенно преобразовывается в третичный фолликул. Третичный фолликул. Имеет сложноустроенную стенку, содержит овоцит первого порядка. Стенка состоит из 2 частей: А. Многослойный фолликулярный эпителий — зернистый слой (гранулеза). Располагается на хорошо выраженной базальной мембране (стекловидная мембрана Славянского). Б. Соединительнотканная часть – тека (покрышка). В зрелом фолликуле 2 слоя: o внутренний рыхлый (большое количество кровеносных сосудов, особые гормонально-активные клетки – текоциты (разновидность интерстициальных клеток), продуцирующие эстрогены являются источником образования опухолей; o фиброзный слой (плотный) состоит из волокон, полость фолликула заполнена; фолликулярной жидкостью, которая содержит эстрогены, гонадокринин (гормон белковой природы, синтезируется фолликулярными клетками, ответственен за атрезию фолликула). На одном из полюсов есть яйценосный холмик, на котором лежит овоцит первого порядка, окруженный лучистым венцом. При образовании ЛГ происходит разрыв фоллликула и выход из яичника половой клетки –овуляция. Половая клетка устремляется в яйцевод, где происходит деление и созревание ее, на месте лопнувшего фолликула происходит образование желтого тела. Его клетки продуцируют прогестерон. Различают 2 вида желтых тел –менструальное и желтое тело беременности. Ментструальное тело меньших размеров (1-2 см в диаметре, в то время как желтое тело беременности 5-6 см), продолжительность жизни его меньше (5-6 дней против нескольких месяцев). 4 стадии развития желтого тела: 1 стадия связана с пролиферацией и делением текоцитов – васкуляризация. 2 стадия железистого превращения. Клетки зернистого слоя и текоциты превращаются в клетки — лютеиноциты, продуцирующие другой гормон. Цитоплазма содержит желтый пигмент. 3 стадия расцвета. Желтое тело достигает максимального размера, максимально много продуцируемых гормонов. 4 стадия — стадия обратного развития. Связана с гибелью железистых клеток. На их месте образуется соединительно-тканный рубец — белое тело, которое со временем рассасывается. Помимо прогестерона, клетки желтого тела синтезируют в небольших количествах эстрогены, андрогены, окситоцин, релаксин. Прогестерон тормозит образование ФСГ и созревание нового фолликула в яичнике, влияет на слизистую матки и молочную железу. Не все фолликулы достигают 4-й стадии развития. Гибель фолликулов 1-й и 2-й стадии проходит незаметно. При гибели фолликулов 3-й и 4-й стадии образуется атретический фолликул. Под влиянием гонадокринина в случае атрезии фолликула сначала гибнет овоцит первого порядка, а затем фолликулярные клетки. От овоцита образуется прозрачная оболочка, которая сливается со стекловидной мембраной и находится в центре атретического фолликула. Интерстициальные клетки активно пролиферируют, их образуется большое количество и образуется атретическое тело (интерстициальная железа). Продуцируют эстрогены. Биологический смысл – предотвращение явлений гиперовуляции, достигается определенный фон в крови эстрогенов перед моментом полового созревания. Все преобразования в фолликуле называются овариальный цикл. Он происходит под влиянием гормонов в 2 фазы: o фолликулиновая фаза под влиянием ФСГ; o лютеиновая фаза под влиянием ЛГ, ЛТГ. Изменения в яичниках вызывают изменения в других органах женской половой системы — яйцеводах, матке, влагалище, молочных железах. Гистология яичника В яичнике (цветн. рис. 2—5) различают: 1) эпителиальную оболочку, 2) белочную оболочку, 3) корковую зону и 4) мозговую зону. 1. Эпителий, покрывающий поверхность свободного края яичника (низкий однослойный цилиндрический), называют «зародышевым», «зачатковым», «герминативным». Термины эти не отвечают функции эпителия яичника взрослой женщины. Правильнее эпителий яичника называть «покровным», «яичниковым». 2. Белочная оболочка, которую покрывает яичниковый эпителий, состоит из плотной соединительной ткани с примесью эластических и гладких мышечных волокон. 3. Корковая зона яичника лежит непосредственно под белочной оболочкой. Паренхима коркового слоя содержит в большом количестве первичные (примордиальные) фолликулы, граафовы (яйценосные) фолликулы в различных стадиях созревания, желтые тела и атретические фолликулы (corpora albicantia). Строение этих образований — см. Менструальный цикл. 4. Мозговая (сосудистая) зона яичника богато снабжена кровеносными сосудами и нервами; эта зона переходит в ворота яичника (hilus ovarii), где часто встречается сеть эпителиальных тяжей и трубочек. Рис. 2. Яичник (малое увеличение): 1 — примордиальные фолликулы в корковом веществе; 2 — граафов пузырек; 3 — мозговое вещество. Рис. 3. Граафов пузырек (большое увеличение): 1 — theca folliculi; 2 — stratum granulosum; 3 — cumulus oophorus; 4 — corona radiata. Рис. 4. Corpus atreticum: 1 — центральное ядро с остатками zona pellucida; 2 — слой железистых клеток. Рис. 5. Corpus luteum: 1 — соединительнотканное ядро; 2 — лютеиновые клетки.
55 Понятие о цитологии гитологии эмбриологии Гистология (от греч. histos — ткань, logos — учение) — наука о строении, развитии и жизнедеятельности тканей животных организмов. Ткани представляют собой систему клеток и неклеточных структур, объединившихся и специализировавшихся в процессе эволюции для выполнения важнейших функций в организме. Для каждой из основных тканевых систем характерны присущие именно им особенности строения, развития и жизнедеятельности. Предметом общей гистологии, или собственно учения о тканях, являются общие закономерности, присущие тканевому уровню организации и отличительные особенности конкретных тканей; предметом частной гистологии — закономерности жизнедеятельности и взаимодействия различных тканей в органах на более высоких уровнях организации. Частная гистология служит основой для изучения микроскопического строения морфофункциональных единиц органов и органов в целом. Курс гистологии включает в себя также цитологию — учение о клетке и эмбриологию — учение о зародыше. Эти самостоятельные курсы предшествуют общей и частной гистологии. Цитология (от греч. kytos — клетка, logos — учение) — наука о развитии, строении и жизнедеятельности клеток. Цитология составляет необходимую часть гистологии, так как клетки являются основой развития и строения тканей. Цитология в последние годы обогатилась многими научными открытиями, внесшими существенный вклад в развитие биологических и медицинских наук и практику здравоохранения. Новые данные о структуре ядра, его хромосомного аппарата легли в основу цитодиагностики наследственных заболеваний, опухолей, болезней крови и многих других болезней. Раскрытие особенностей ультраструктуры и химического состава клеточных мембран является основой для понимания закономерностей взаимодействия клеток в тканевых системах, защитных реакциях и др. В медицинской практике широко используется цитодиагностика. Клетки здорового и больного организма изучаются в мазках крови и костного мозга, спинномозговой жидкости, слюны, мочи, в образцах различных органов, взятых при биопсии. Эмбриология (от греч. embryon — зародыш, logos — учение) — учение о зародыше, о закономерностях его развития. В курсе эмбриологии, преподаваемом в медицинском вузе, основное внимание обращается на закономерности эмбрионального развития человека. Особое внимание в обращается на источники развития и механизмы образования тканей (т.е. гистогенез) на определенном этапе эмбриогенеза. Закономерности гистогенеза определяют морфофункциональные особенности тканевых структур в постнатальном развитии, в частности их способность к регенерации. Поэтому изучение основных этапов эмбрионального развития предшествует изучению тканей. Таким образом, объединение гистологии, цитологии и эмбриологии в один предмет не формально, а отражает внутренние естественные связи между ними. Гистология с цитологией и эмбриологией, как и другие фундаментальные биологические науки, решает главную задачу — выяснение структурной организации процессов жизнедеятельности и в связи с этим — возможности целенаправленного воздействия на них.
57 Аксоны и Дендриты. Нервные волокна. Аксон — обычно длинный отросток, приспособленный для проведения возбуждения и информации от тела нейрона или от нейрона к исполнительному органу. Дендриты — как правило, короткие и сильно разветвлённые отростки, служащие главным местом образования влияющих на нейрон возбуждающих и тормозных синапсов (разные нейроны имеют различное соотношение длины аксона и дендритов), и которые передают возбуждение к телу нейрона. Нейрон может иметь несколько дендритов и обычно только один аксон. Один нейрон может иметь связи со многими (до 20-и тысяч) другими нейронами. В центральной нервной системе оболочки отростков нейронов образуются отростками олигодендроглиоцитов, а в периферической —нейролеммоцитами Шванна. Безмиелиновые нервные волокна находятся преимущественно в составе автономной, или вегетативной, нервной системы. Нейролеммоциты оболочек безмиелиновых нервных волокон, располагаясь плотно, образуют тяжи. В нервных волокнах внутренних органов, как правило, в таком тяже имеется не один, а несколько осевых цилиндров, принадлежащих различным нейронам. Они могут, покидая одно волокно, переходить в соседнее. Такие волокна, содержащие несколько осевых цилиндров, называются волокнами кабельного типа. По мере погружения осевых цилиндров в тяж нейролеммоцитов оболочки последних прогибаются, плотно охватывают осевые цилиндры и, смыкаясь над ними, образуют глубокие складки, на дне которых и располагаются отдельные осевые цилиндры. Сближенные в области складки участки оболочки нейролеммоцита образуют сдвоенную мембрану — мезаксон, на которой как бы подвешен осевой цилиндр. Миелиновые нервные волокна встречаются как в центральной, так и в периферической нервной системе. Они значительно толще безмиелиновых нервных волокон. Они также состоят из осевого цилиндра, «одетого» оболочкой из нейролеммоцитов Шванна, но диаметр осевых цилиндров этого типа волокон значительно толще, а оболочка сложнее. Миелиновый слой оболочки такого волокна содержит значительное количество липидов, поэтому при обработке осмиевой кислотой он окрашивается в темно-коричневый цвет. В миелиновом слое периодически встречаются узкие светлые линии—насечки миелина, или насечки Шмидта — Лантермана. Через определенные интервалы (1—2 мм) видны участки волокна, лишенные миелинового слоя, — это т.н. узловатые перехваты, илиперехваты Ранвье.
57 Аксоны и Дендриты. Нервные волокна. Аксон — обычно длинный отросток, приспособленный для проведения возбуждения и информации от тела нейрона или от нейрона к исполнительному органу. Дендриты — как правило, короткие и сильно разветвлённые отростки, служащие главным местом шен осевой цилиндр. Миелиновые нервные волокна встречаются как в центральной, так и в периферической нервной системе. Они значительно толще безмиелиновых нервных волокон. Они также состоят из осевого цилиндра, «одетого» оболочкой из нейролеммоцитов Шванна, но диаметр осевых цилиндров этого типа волокон значительно толще, а оболочка сложнее. Миелиновый слой оболочки такого волокна содержит значительное количество липидов, поэтому при обработке осмиевой кислотой он окрашивается в темно-коричневый цвет. В миелиновом слое периодически встречаются узкие светлые линии—насечки миелина, или насечки Шмидта — Лантермана. Через определенные интервалы (1—2 мм) видны участки волокна, лишенные миелинового слоя, — это т.н. узловатые перехваты, илиперехваты Ранвье.
58 Различие сперматогенеза и овогенеза 1. Мы выяснили, что гаметогенез включает стадии размножения, роста и созревания клеток. Сперматогенез включает также стадию формирования (ее нет при овогенезе), в этом заключаются отличия сперматогенеза от овогенеза. Сперматозоиды проходят дополнительную четвертую стадию для того, чтобы приобрести своеобразную форму и сформировать аппарат движения.2. Второе отличие сперматогенеза от овогенеза: из сперматоцита I порядка получается четыре половых клетки, а из ооцита I порядка получается одна полноценная половая клетка.3. Яйцеклетки образуются циклически, процесс повторяется через 21-35 дней (менструальный цикл). После гибели яйцеклетки, что сопровождается кровотечением, изменившийся гормональный фон дает толчок к созреванию другой яйцеклетки. Сравнительная характеристика овогенеза и сперматогенеза показывает, что у женщин мейоз начинается в период внутриутробного развития. Ооциты I порядка у новорожденной девочки останавливаются в фазе мейоз I, и завершается созревание ооцита к моменту полового созревания. У мальчиков процесс образования сперматозоидов идет непрерывно и сохраняется в течение всей половой зрелости мужчины.4. Из характеристики овогенеза и сперматогенеза следует, что существуют значительные различия в количестве образованных половых клеток в женском и мужском организме. Взрослый мужчина производит 30 миллионов спермиев в день, а женщина - порядка 500 зрелых яйцеклеток за всю свою жизнь.5. Различия сперматогенеза и овогенеза заключаются также в том, что стадия размножения при сперматогенезе идет постоянно, а при овогенезе заканчивается после рождения.6. Стадия роста при сперматогенезе короче, чем при овогенезе7. Стадия созревания при овогенезе имеет особенности, которые заключаются в неравномерности делений при созревании, что приводит к выделению полярных телец, что отсутствует при сперматогенезе.8. Различия сперматогенеза и овогенеза заключаются в том, что сперматогенез более подвержен влиянию внешней среды, нежели овогенез, что связано с различием в расположении половых органов - семенники находятся вне брюшной полости.9. Из сравнительной характеристики овогенеза и сперматогенеза можно увидеть, что, поскольку образование яйцеклеток начинается еще до рождения девочки, а завершается для яйцеклетки только после ее оплодотворения, то неблагоприятные факторы внешней среды могут повлечь генетические аномалии у потомства.
59 Гистологическое строение легких. Легкое - компактный (паренхиматозный) орган, снаружи одетый серозной оболочкой - легочной плеврой. Соединительнотканная строма образует тонкую, но плотную капсулу, в составе которой много эластических волокон. Междольковые прослойки делят орган на дольки, заметные невооруженным глазом. В них проходят бронхи, нервы, кровеносные сосуды. Дольчатость легкого лучше выражена у крупного рогатого скота и свиньи. Внутридольковые прослойки представлены в основном фибробластами и эластическими волокнами. В состав паренхимы легкого входят воздухоносные пути - бронхиальное дерево и респираторные отделы - альвеолярное дерево. Бронхиальное дерево состоит из бронхов разного калибра: крупных, средних, мелких и бронхиол. Строение стенки бронхов меняется с уменьшением их диаметра. Стенка главного бронха отличается от стенки трахеи тем, что в слизистой оболочке его появляются пучки гладкомышечных клеток, хрящевые кольца фиброзно-хрящевой оболочки замкнутые. В стенке крупных бронхов заметна мышечная пластинка слизистой оболочки. При сокращении ее слизистая оболочка собирается в мелкие складки. Хрящевые кольца имеют вид хрящевых пластинок неправильной формы. Адвентиция переходит в междольковую соединительную ткань Средний бронх а на препарате заметен невооруженным глазом в виде маленького сиреневатого кольца. Слизистая оболочка его выстлана многорядным мерцательным эпителием 1, но клетки ниже, чем в трахее. Бокаловидные клетки встречаются реже. В слизистой оболочке хорошо развиты эластические волокна и мышечная пластинка 2, в результате чего слизистая оболочка собрана в складки. В подслизистой основе видны концевые отделы серозно-слизистых желез 3 в большем количестве, чем в трахее. Фиброзно-хрящевая оболочка тонкая, содержит мелкие островки хряща 4. Вместо адвентиции - междольковая соединительная ткань 5. Стенка малого бронха б состоит только из слизистой оболочки, покрытой реснитчатым эпителием с редуцированным количеством слоев. Чем мельче бронх, тем меньше слоев эпителия и ниже клетки. Мышечная пластинка 2 тонкая, железы и хрящ отсутствуют. Наружной оболочкой является рыхлая соединительная ткань. Конечный участок воздухоносных путей - концевая (терминальная) бронхиола в выстлана однослойным кубическим реснитчатым эпителием. В собственной пластинке ее (очень небольшой толщины) расположены эластические волокна и отдельные пучки гладких мышечных клеток. Респираторные отделы имеют вид кружева; занимают основную площадь препарата. Состоят они из альвеол, альвеолярных ходов и дыхательных (респираторных) бронхиол, объединенных в ацинусы. Ацинус - это участок альвеолярного дерева, отходящий от одной терминальной бронхиолы. Дольку составляют 10-20 ацинусов. Терминальная бронхиола распадается на несколько респираторных бронхиол г, эпителий которых утратил реснички, но в собственной пластинке слизистой оболочки еще имеются отдельные пучки гладких мышечных клеток. Респираторные бронхиолы разделяются на альвеолярные ходы д, каждый из которых кончается двумя альвеолярными мешочками е. Стенка их образована альвеолами. Альвеола ж имеет вид пузырька, открытого внутрь мешочка, или хода. Внутренняя поверхность альвеолы Б выстлана однослойным плоским эпителием 6, находящимся на базальной мембране 7. Снаружи к базальной мембране вплотную прилежат кровеносные капилляры 8, проходящие по чрезвычайно тонким межальвеолярным перегородкам, содержащим эластические волокна 9. Местами отсутствует и этот слой, и граница между воздухом и кровью состоит только из альвеолярной клетки, базальной мембраны и эндотелиальной клетки. Причем ядра этих клеток лежат напротив друг друга, а безъядерные участки цитоплазмы вместе с базальной мембраной имеют толщину около 0,5 мкм. Этот барьер между кровью и воздухом не имеет пор. Давление кислорода в венозной крови, поступающей в легочные капилляры, гораздо ниже, а углекислого газа - гораздо выше, чем в альвеолярном воздухе. Разность давлений и обеспечивает диффузию кислорода из альвеолярного воздуха в кровь, углекислого газа - в обратном направлении.
60 гистологическое строение Рыхлая волокнистая соединительная ткань, клеточный составОсновными клетками соединительной ткани являются фибробласты (семейство фибриллообразующих клеток), макрофаги, тучные клетки, адвентициальные клетки, плазматические клетки, перициты, жировые клетки, а также лейкоциты, мигрирующие из крови; иногда встречаются пигментные клетки.Фибробласты (фибробластоциты) (от лат. fibra — волокно, греч. blastos — росток, зачаток) — клетки, синтезирующие компоненты межклеточного вещества: белки (например, коллаген, эластин), протеогликаны, гликопротеины.В эмбриональном периоде ряд мезенхимных клеток зародыша дают начало дифферону фибробластов, к которому относят:стволовые клетки,полустволовые клетки-предшественники,малоспециализированные фибробласты,дифференцированные фибробласты (зрелые, активно функционирующие),фиброциты (дефинитивные формы клеток),миофибробласты и фиброкласты.С главной функцией фибробластов связаны образование основного вещества и волокон (что ярко проявляется, например, при заживлении ран, развитии рубцовой ткани, образовании соединительнотканной капсулы вокруг инородного тела).Дифференцированные зрелые фибробласты крупнее по размеру. Это активно функционирующие клетки.В зрелых фибробластах осуществляется интенсивно биосинтез коллагеновых, эластиновых белков, протеогликанов, которые необходимы для формирования основного вещества и волокон. Эти процессы усиливаются в условиях пониженной концентрации кислорода. Стимулирующими факторами биосинтеза коллагена являются также ионы железа, меди, хрома, аскорбиновая кислота. Один из гидролитических ферментов — коллагеназа — расщепляет внутри клеток незрелый коллаген, что регулирует на клеточном уровне интенсивность секреции коллагена.Фибробласты – это подвижные клетки. В их цитоплазме, особенно в периферическом слое, располагаются микрофиламенты, содержащие белки типа актина и миозина. Движение фибробластов становится возможным только после их связывания с опорными фибриллярными структурами с помощью фибронектина — гликопротеина, синтезируемого фибробластами и другими клетками, обеспечивающего адгезию клеток и неклеточных структур. Во время движения фибробласт уплощается, а его поверхность может увеличиться в 10 раз. Плазмолемма фибробластов является важной рецепторной зоной, которая опосредует воздействие различных регуляторных факторов. Активизация фибробластов обычно сопровождается накоплением гликогена и повышенной активностью гидролитических ферментов. Энергия, образуемая при метаболизме гликогена, используется для синтеза полипептидов и других компонентов, секретируемых клеткой. По способности синтезировать фибриллярные белки к семейству фибробластов можно отнести ретикулярные клетки ретикулярной соединительной ткани кроветворных органов, а также хондробласты и остеобласты скелетной разновидности соединительной ткани.Фиброциты — дефинитивные (конечные) формы развития фибробластов. Эти клетки веретенообразные с крыловидными отростками. [Они содержат небольшое число органелл, вакуолей, липидов и гликогена.] Синтез коллагена и других веществ в фиброцитах резко снижен. Миофибробласты — клетки, сходные с фибробластами, сочетающие в себе способность к синтезу не только коллагеновых, но и сократительных белков в значительном количестве. Фибробласты могут превращаться в миофибробласты, функционально сходные с гладкими мышечными клетками, но в отличие от последних имеют хорошо развитую эндоплазматическую сеть. Такие клетки наблюдаются в грануляционной ткани заживающих ран и в матке при развитии беременности.Фиброкласты — клетки с высокой фагоцитарной и гидролитической активностью, принимают участие в «рассасывании» межклеточного вещества в период инволюции органов (например, в матке после окончания беременности). Они сочетают в себе структурные признаки фибриллообразующих клеток (развитую гранулярную эндоплазматическую сеть, аппарат Гольджи, относительно крупные, но немногочисленные митохондрии), а также лизосомы с характерными для них, гидролитическими ферментами. Выделяемый ими за пределы клетки комплекс ферментов расщепляет цементирующую субстанцию коллагеновых волокон, после чего происходят фагоцитоз и внутриклеточное переваривание коллагена.Следующие клетки волокнистой соединительной ткани уже не относятся к дифферону фибробластов. Макрофаги (или макрофагоциты) (от греч. makros — большой, длинный, fagos — пожирающий) — это гетерогенная специализированная клеточная популяция защитной системы организма.
61 Классификация эпителиев по происхождениюЭпителиальные ткани развиваются из всех трех зародышевых листков. Эктодермальный эпителий покрывает тело снаружи и входит в состав кожи, начальных участков дыхательных путей, роговицы глаза, переднего и заднего отделов кишечной трубки. Энтодермальный эпителий выстилает изнутри пищеварительную трубку и является основной составной частью пищеварительных желез и органов дыхания. Мезодермальный эпителий образует внутреннюю выстилку полостей тела, входит в состав серозных оболочек (мезотелий), половых желез, матки, почек и мочевыводящих путей. Основные признаки эпителиальных тканей: 1) пограничное положение, в результате чего эпителий отграничивает организм от внешней или внутренней среды и одновременно осуществляет с ней связь; 2) чисто клеточное строение, отсутствие межклеточного вещества; 3) расположение клеток как в виде сплошного пласта, что является необходимым условием функционирования эпителия, так и на базальной мембране, образованной деятельностью клеток эпителия и соединительной ткани; 4) морфологическая и функциональная полярность клеток, которая выражается в том, что апикальная (верхняя) и базальная (нижняя) части клетки различаются по структуре и функции, а в многослойных эпителиях, кроме того, отличаются и клетки, находящиеся в разных слоях.Эпителиальные ткани можно классифицировать по структурным и функциональным признакам. В связи с преобладанием той или иной функции эпителий делят на покровный (защитная функция), всасывающий (трофическая функция), почечный (выделительная функция), железистый (секреторная функция), эпендимоглиальный (слуховой, вкусовой - участие в восприятии раздражения) и др. Морфологическая классификация эпителиев основана на количестве слоев и форме клеток. Принято различать эпителий однослойный, который делится на однорядный (плоский, кубический, призматический) и многорядный (мерцательный), и многослойный - ороговевающий, неороговевающий и переходный. В рамках этой классификации эпителий делят на покровный (одевает организм и органы снаружи), выстилающий (лежит со стороны полости органа) и паренхиматозный (образует основную ткань компактных, паренхиматозных органов), в том числе и желез железистый эпителий).
63. Круги кровообращения. Типы сосудов,Сердце играет роль мышечного насоса, обеспечивающего ритмическое поступление крови в систему сосудов. Это достигается мощным развитием специальной сердечной мускулатуры и наличием особых клеток - водителей ритма.Крупные артерии вблизи сердца растягиваются при поступлении порции крови из сердца и возвращаются к прежним размерам, выбрасывая кровь в дальнейшие участки сосудистого русла. Благодаря этому кровоток остается непрерывным. Эта функция обеспечивается мощным развитием эластических элементов в стенке таких сосудов.Средние и мелкие артерии приносят кровь к различным органам и их частям, регулируя кровоток в зависимости от функционального состояния этих органов. Это обеспечивается развитыми мышечными элементами в стенке таких артерий. В связи с тем, что кровь в артериях течет под большим давлением, их стенка имеет большую толщину и содержит развитые эластические элементы.Артериолы — самые мелкие артерии. В артериолах происходит резкий перепад давления, — от высокого в артериях до низкого в капиллярах. Это обусловлено значительным количеством этих сосудов, их узким просветом и наличием мышечных элементов в стенке. Общее давление в артериальной системе определяется в значительной степени тонусом именно артериол. Капилляры осуществляют двусторонний обмен веществ между кровью и тканями, что достигается благодаря их огромной общей поверхности и тонкой стенке.Венулы собирают из капилляров кровь, которая движется под низким давлением. Их стенки тонкие, что также способствует обмену веществ и облегчает миграцию клеток из крови в ткани.Вены обеспечивают возврат крови к сердцу. Они характеризуются широким просветом, тонкой стенкой со слабым развитием эластических и мышечных элементов. В венах имеются клапаны, препятствующие обратному току крови.Однослойный плоскийэпителий, выстилающий изнутри сердце, кровеносные и лимфатические сосуды, имеет собственное название — эндотелий. Его клетки — эндотелиоциты — имеют полигональную форму, обычно удлиненную по ходу сосуда, и связаны друг с другом плотными и щелевыми контактами.Стенка сосудов состоит из трех оболочек: внутренней оболочки - интимы (tunica interna s. intima);средней оболочки - медии (tunica media);наружной оболочки - адвентиции (tunica externa s. adventitia).Их толщина, тканевый состав и функциональные особенности неодинаковы в сосудах разных типов.Внутренняя оболочка (интима) образована:эндотелием (разновидностью плоского однослойного эпителия); подэндотелиальным слоем, состоящим из рыхлой соединительной ткани;внутренней эластической мембраной.Средняя оболочка (медия) включает слои циркулярно расположенных гладкомышечных клеток, а также сеть коллагеновых, ретикулярных и эластических волокон.Наружная оболочка (адвентиция) образована:наружной эластической мембраной, которая может быть представлена лишь отдельными волокнами; рыхлой волокнистой соединительной тканью, содержащей нервы и сосуды, питающие собственную стенку сосудов - нервы сосудов и сосуды сосудов.Круги кровообращения.В кровеносной системе различают два круга кровообращения. В малом, легочном (дыхательном), круге кровообращения кровь идет от правого желудочка сердца по легочному стволу (венозная), который делится на две легочные артерии-для каждого легкого. В легких артерии ветвятся до капилляров альвеол, где происходит газообмен. Капилляры собираются в легочные вены (артериальная кровь), впадающие в левое предсердие. В большом, системном (трофическом), круге кровообращения кровь идет от левого желудочка в аорту, откуда она разносится по всему телу. Из органов и тканей тела кровь собирается в краниальную и каудальную полые вены, непарную вену, открывающиеся в правое предсердие.
74. принципы строения трубкообразных органов на примере пищевода ТРУБКООБРАЗНЫЕ органы – служат путями для прохождения определенного содержимого, т. е. являются в организме каналами или трактами.Стенка трубкообразного органа состоит из трех оболочек: слизистой, мышечной и серозной.1. Слизистая оболочка (tunica mucosa) – выстилает внутреннюю поверхность полых органов, влажная, блестящая, покрыта слизью (муцином). Имеет довольно сложное строение и включает обязательные и необязательные компоненты: а) эпителиальный слой – покрывает слизистую со стороны просвета и имеет различное строение (в ротовой полости и пищеводе – многослойный плоский, в дыхательной трубке - мерцательный) б) собственно слизистая оболочка – представлена соединительной тканью, с подходящими к ней сосудами и нервами. Этот слой отделен от эпителиального базальной мембраной.в) подслизистый слой - необязательный компонент слизистой оболочки, наиболее выражен в органах, способных при наполнении сильно расширяться (кишечник, желудок) и отсутствует там, где слизистая должна плотно соединяться с подлежащим слоем (десны, язык). г) мышечный слой слизистой олочки – благодаря наличию гладкомышечных элементов позволяет слизистой собираться в складки (продольные – в пищеводе, спиралевидные – в сычуге и т. д.), что дает возможность ей растягиваться.2. Мышечная оболочка – (tunica muscularis) средний слой стенки трубкообразного органа, представлен гладкой мышечной тканью, волокна которой формируют два пласта – продольный и циркулярный. Циркулярный слой сокращаясь, изменяет диаметр трубки, а продольный - длину. Поочередное сокращение пластов приводит к перистальтике. Там, где трубка крепится к костям, гладкая мышечная ткань заменяется на поперечно-полосатую. В некоторых органах этот слой может отсутствовать (твердое небо). В органах, где просвет трубки должен постоянно зиять мышечная ткань заменяется на хрящевую или фиброзную (дыхательная трубка).3. Серозная оболочка (tunica serosa) – наружный слой трубкообразного органа, представлен одним слоем плоских клеток – мезотелем, которые выделяют небольшое количество серозной жидкости, придавая органу гладкий и блестящий вид. Серозная оболочка развивается в тех случаях, когда трубкообразный орган лежит в серозной полости (грудной, брюшной, тазовой), если орган располается вне полости – наружная оболочка – адвентиция, представленная рыхлой соединительной тканью, которая, окружая трубку, прикрепляет его к окружающим тканям (шейная часть пищевода).В толще стенки трубчатых органов или снаружи от трубки находятся железы, выделяющие специфическое вещество (слюну, слизь, желчь). Железы, располагающиеся в основе слизистой оболочки, называются пристенными. Они могут быть одноклечные или многоклеточные, обычно такие железы не видны невооруженным глазом. Железы, лежащие за пределами стенки трубчатого органа, в который они изливают свой секрет, посредством протоков называются застенными (околоушная слюнная железа, печень, поджелудочная железа).Во всех трубчатых органах много кровеносных и лимфатических сосудов. Они ветвятся в подслизистом слое, а в компактном органе – в строме. Иннервируются внутренние органы вегетативной нервной системой. .
75. гистологическое строение центральной нервной системы Нервная система имеет довольно сложное гистологическое строение. В ее состав входят нейроны (нервные клетки), нервные волокна, нейроглия и соединительнотканные элементы. Нейроны являются основной структурно-функциональной частью нервной системы (. 3). Величина и форма нейрона зависят от длины и количества его отростков. Отростки принято делить на дендриты (цитоплазматические) и аксоны, или нейриты (осевоцилиндрические). Дендриты, древовидно разветвляющиеся выросты цитоплазмы нейрона, проводят раздражение в сторону тела нейрона. Аксон представляет собой осевоцилиндрический длинный одиночный отросток, несущий импульсы от нервной клетки; он оканчивается короткими разветвлениями.Тело нейрона состоит из цитоплазмы и ядра. В цитоплазме имеются хроматофильное (тигроидное) вещество и нейрофибриллы, являющиеся специфическими компонентами нервных клеток. Хроматофильное вещество имеется в теле нейрона и крупных дендритах в виде глыбок различной величины. Оно изменяется при различных функциональных состояниях нейрона, поэтому его исследование может иметь определенное диагностическое значение. Нейрофибриллы в виде тончайших нитей находятся а теле клетки и ее отростках. Кроме того, нейрон содержит сетчатый аппарат Гольджи, митохондрии и другие органоиды, а также входящее в ядро ядрышко, богатое нуклеиновыми кислотами.Нервные волокна образованы отростками нейронов. В зависимости от того, имеют ли они слой миелина, они делятся на безмиелиновые (безмякотные) и миелиновые (мякотные). Первые преобладают в вегетативной, а вторые—в соматической части нервной системы.Миелиновое нервное волокно состоит из осевого цилиндра, являющегося продолжением аксона нейрона, слоя миелина и слоя нейролеммоцитов — неврилеммы (шванновской оболочки) (. 4).Осевой цилиндр состоит из аксоплазмы и нейрофибрилл, играющих важную роль в регенерации нервного волокна. При повреждении осевого цилиндра аксоплазма начинает вытекать из конца центрального отрезка, вследствие чего образуются ее натеки, в которые врастают нейрофибриллы. Слой миелина (миелиновая, или мякотная, оболочка), окутывающий осевой цилиндр, имеет значительную толщину на протяжении всего нервного волокна и содержит большое количество миелина, состоящего из жироподобных веществ. Истончаясь в отдельных участках, он образует межсег-ментарные узлы нервного волокна (перехваты Ранвье). В слое миелина также имеются глубокие воронкообразные впадины, видимые на продольных срезах как канальцы, достигающие осевых цилиндров в косом направлении,— насечки миелина (насечки Шмидта—Лантермана).Цитоплазма нейролеммоцитов (шванновских клеток), выпячиваясь в такие воронки, образует отростки, которые также достигают осевых цилиндров. Благодаря таким отросткам нейро-леммоциты, как полагает Б. С. Дойников, осуществляют регуляцию трофических процессов в осевом цилиндре нервного волокна. Отличительной чертой нервных волокон в центральной нервной системе является то, что они не имеют узлов нервного волокна и нейролеммы и осевые цилиндры сопровождаются глиальными элементами.Нейролеммоциты выполняют роль периферической глии. Наружный слой нейролеммоцита с его ядром и большей частью цитоплазмы составляет нейролемма. Внутренний слой цитоплазмы содержит мнелиновые пластинки. При повреждении нервного волокна нейролеммоциты пролиферируют и образуют мостики (бюнгеровские ленты) между его центральным и периферическим отрезками, играя таким образом важную роль в регенерации.Нейроглия играет роль соединительной ткани центральной нервной системы (. 5). Различают макро- и микроглию. В свою очередь макроглия состоит из астроцитной глии и олигодендроглииАстроцитная г л и я—синцитиальное образование, состоящее из крупных многоотростчатых астроцитов. Отростки одних астроцитов, соединяясь с отростками других астроцитов, образуют балки, между которыми находятся нервные клетки.Олигодендроглия выполняет опорную функцию преимущественно для отростков нейронов, сопровождая их на всем протяжении до концевых аппаратов. В периферической нервной системе она, как отмечалось выше, представлена нейролеммоцитами. Микроглия состоит из мелких отросчатых клеток, выполняющих защитную функцию. Помимо опорной глии пуща и защитная функция. Клетки глии принимают участие в разнообразных реакциях в ответ на действие вредных факторов на нервную систему. При этом клетки глии сначала увеличиваются в объеме, затем размножаются, астроциты и клетки олигодендроглии замещают гибнущую нервную ткань в виде глиозного рубца. Наряду с регенеративными процессами глия может в патологических условиях подвергаться и дегенеративным изменениям. Некоторые авторы приписывают нейроглии внутекреторную функцию, а также участие в процессах хранения информации.
76. строение амниона хориона и их функции ветеринарияАмнион,Или амниотическая оболочка, обеспечивает образование водной среды (амниотической жидкости), в которой происходит развитие зародыша, осуществляет экстраплацентарную гуморальную связь между организмами матери и плода. Эволюционно амнион возник в процессе выхода животных на сушу. В эмбриогенезе он появляется в первую фазу гаструляции почти одновременно с желточным мешком в виде амниотического пузырька, локализующегося над эмбриональным диском, в связи с чем его дном является эпибласт. Одним из своих участков амниотический пузырек прикрепляется к мезодерме, выстилающей изнутри хориальную оболочку. Здесь формируется так называемая амниотическая, или зародышевая, ножка, в будущем преобразующаяся в пупочный канатик.Стенка амниотического пузырька образована двумя слоями: внезародышевой эктодермой и прилежащей к ней снаружи внезародышевой мезодермой, являющейся продолжением париетального листка спланхнотома.Внезародышевая эктодерма является источником развития амниотического однослойного эпителия, который выполняет как секреторную (в области плацентомов), так и резорбционную (в остальных зонах амниона) функции. Внезародышевая мезодерма дает начало мезенхиме, из которой развивается внезародышевая соединительная ткань стенки амниона, которая образует 2 слоя. Один из них, непосредственно прилежащий к базальной мембране амниотического эпителия, представлен плотной волокнистой соединительной тканью, а другой, наружный, - образован рыхлой слизистой соединительной тканью (губчатый слой), состоящей из небольшого количества коллагеновых волокон и кислых гликозаминогликанов (ГАГ). По мере роста зародыша амниотический пузырь быстро увеличивается в размерах и уже вскоре окружает все его тело. Вследствие секреторной деятельности амниотического эпителия полость пузыря заполняется жидкостью, в результате чего зародыш оказывается полностью в ней погруженным. Между губчатым слоем амниона и соединительнотканной основой хориальной оболочки находится амнио-хориальное пространство, которое по мере увеличения размеров амниотического пузыря уменьшается до минимума и губчатый слой местами соединяется со стенкой хориона. В области амниотической ножки он прочно с нею срастается, в результате чего формирующийся в дальнейшем из амниотической ножки пупочный канатик оказывается снаружи покрытым амниотическим эпителием.Основная функция амниона - выработка околоплодных вод, являющихся средой для развития зародыша, которая защищает его от механических повреждений. Кроме того, амнион участвует в удалении продуктов метаболизма плода, а также в поддержании необходимого состава и концентрации электролитов, кислотно-щелочного равновесия, обеспечивая тем самым гомеостаз. Велика роль амниона и как барьера для вредных веществ.Хорион, или ворсинчатая оболочка, эволюционно появляется у плацентарных млекопитающих. Источником его развития являются трофобласт и внезародышевая париетальная мезодерма. Сначала трофобласт образован одним слоем клеток (бластомеров), снаружи от которых на очень ранних этапах появляется еще один неклеточный слой и, таким образом, трофобласт приобретает двухслойное строение: внутренний его слой клеточный - цитотрофобласт (ЦТ), а наружный - неклеточный - симпластотрофобласт, или синцитиотрофобласт (СТ). При этом СТ происходит из цитотрофобласта вследствие незавершенного митотического деления его клеток (эндомитоз). На поверхности СТ вскоре формируются небольшие выросты - первичные ворсинки, которые вырабатывают ферменты, обладающие высокой протеолитической активностью. Благодаря этому осуществляется разрушение материнских тканей и имплантация зародыша в слизистую оболочку матки (эндометрий), что свойственно для человека и животных с гемохориальным типом плацент.В процессе выселения из эмбрионального диска внезародышевая мезодерма преобразуется в мезенхиму, которая обрастает двухслойный трофобласт изнутри и вместе с ним формирует хорион.В дальнейшем происходят количественные и качественные преобразования: первичные изначально трофобластические ворсинки превращаются во вторичные вследствие врастания в них мезенхимы, очень скоро дифференцирующейся во внезародышевую соединительную ткань. Количество вторичных ворсинок быстро нарастает, а в их соединительнотканной строме начинается васкулогенез и с этого момента ворсинки называются уже третичными (рис. 4). В покрывающем ворсинки СТ усиливается синтез протеолитических ферментов, активно воздействующих на структурные компоненты слизистой оболочки матки, - начинается плацентогенез.
77. Гистологическое строение матки разных видов с\х животных. У всех животных матка
 2015-02-18
2015-02-18 2901
2901 пластинки).
пластинки). 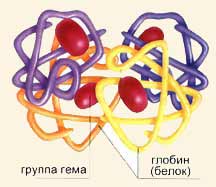 Электронная микрофотография гемолиза эритроцитов и образование их “теней” 1 – дискоцит, 2 – эхиноцит, 3 – “тени” (оболочки) эритроцитов.
Электронная микрофотография гемолиза эритроцитов и образование их “теней” 1 – дискоцит, 2 – эхиноцит, 3 – “тени” (оболочки) эритроцитов. 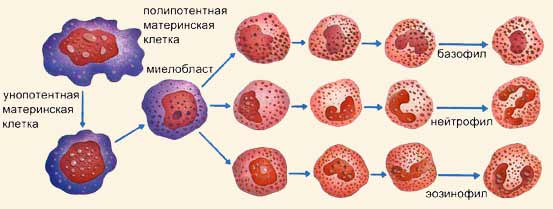 или кровяные пластинки – плоские клетки неправильной округлой формы диаметром 2 – 5 мкм. Тромбоциты человека не имеют ядер - это фрагменты клеток, которые меньше половины эритроцита. Количество тромбоцитов в крови человека составляет 180 – 320х10'/л, или 180 000 – 320 000 в 1 мкл. Имеют место суточные колебания: днем тромбоцитов больше, чем ночью. Увеличение содержания тромбоцитов в периферической крови называется тромбоцитозом, уменьшение – тромбоцитопенией.
или кровяные пластинки – плоские клетки неправильной округлой формы диаметром 2 – 5 мкм. Тромбоциты человека не имеют ядер - это фрагменты клеток, которые меньше половины эритроцита. Количество тромбоцитов в крови человека составляет 180 – 320х10'/л, или 180 000 – 320 000 в 1 мкл. Имеют место суточные колебания: днем тромбоцитов больше, чем ночью. Увеличение содержания тромбоцитов в периферической крови называется тромбоцитозом, уменьшение – тромбоцитопенией.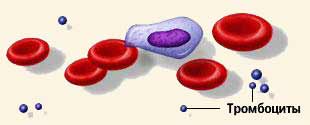
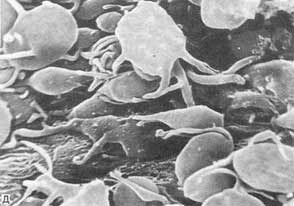 Тромбоциты, прилипшие к стенке аорты в зоне повреждения эндотелиального слоя.
Тромбоциты, прилипшие к стенке аорты в зоне повреждения эндотелиального слоя. 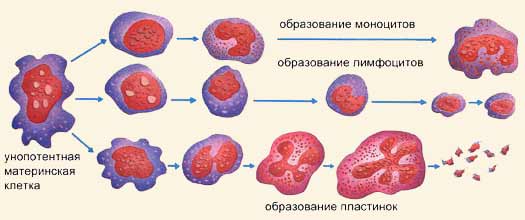
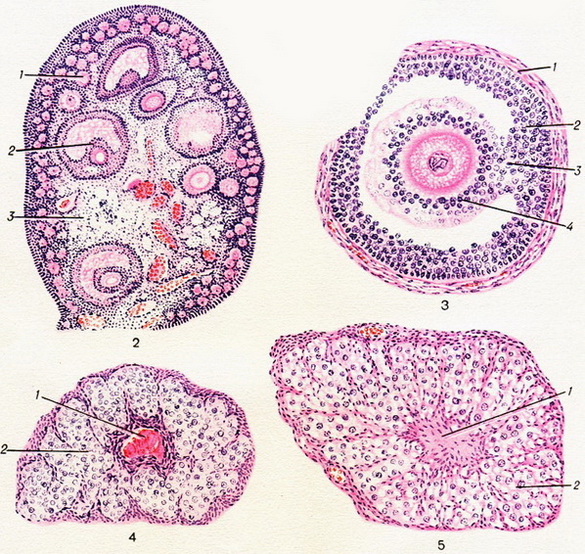 Рис. 2. Яичник (малое увеличение): 1 — примордиальные фолликулы в корковом веществе; 2 — граафов пузырек; 3 — мозговое вещество. Рис. 3. Граафов пузырек (большое увеличение): 1 — theca folliculi; 2 — stratum granulosum; 3 — cumulus oophorus; 4 — corona radiata. Рис. 4. Corpus atreticum: 1 — центральное ядро с остатками zona pellucida; 2 — слой железистых клеток. Рис. 5. Corpus luteum: 1 — соединительнотканное ядро; 2 — лютеиновые клетки.
Рис. 2. Яичник (малое увеличение): 1 — примордиальные фолликулы в корковом веществе; 2 — граафов пузырек; 3 — мозговое вещество. Рис. 3. Граафов пузырек (большое увеличение): 1 — theca folliculi; 2 — stratum granulosum; 3 — cumulus oophorus; 4 — corona radiata. Рис. 4. Corpus atreticum: 1 — центральное ядро с остатками zona pellucida; 2 — слой железистых клеток. Рис. 5. Corpus luteum: 1 — соединительнотканное ядро; 2 — лютеиновые клетки.  8463
8463 7918
7918