 2015-02-24
2015-02-24 11915
119151. Открытие и общие свойства фитогормонов. Работы Ч. Дарвина, Бойсена-Иенсена, Холодного, Вента. Гормональная теория тропизмов.
Фитогормоны — низкомолекулярные органические вещества, вырабатываемые растениями и имеющие регуляторные функции. Гормоны в органах - ауксинами богаче всего верхушечные меристемы стебля, гиббереллинами и флоригеном — листья, цитокининами — корни и созревающие семена. Фитогормоны регулируют многие процессы жизнедеятельности растений: прорастание семян, рост, дифференциацию тканей и органов, цветение, созревание плодов и т. п. Химические соединения, которые вырабатываются в одних частях растений и оказывают своё действие в других, проявляют свой эффект в исключительно малых концентрациях, обладают (в отличие от ферментов) обычно меньшей специфичностью действия на процессы роста и развития, что объясняется разным состоянием работы генов воспринимающих клеток, от которого зависит результат действия гормона, а также разным соотношением между собой различных фитогормонов (гормональным балансом). Эффект фитогормонов в значительной мере определяется действием других внутренних и внешних факторов. Гормоны разных растений могут отличаться по химической структуре, поэтому они сгруппированы на основании их эффекта на физиологию растений и общему химическому строению. Пять групп фитогормонов: ауксины, гиббереллины, цитокинины, абсцизовая кислота, газ этилен. В последнее время к ним относят брассины (брассиностероиды). Условно можно отнести первые три группы—ауксины, гиббереллины и цитокинины и частично брассины — к веществам стимулирующего характера, тогда как абсцизовую кислоту и этилен — к ингибиторам.
Ч. Дарвин описал опыты по изучению изгибания проростков злака по направлению к свету. Было установлено, что свет воспринимается только самой верхушкой колеоптиля, тогда как изгиб происходит в нижележащей зоне, которая сама по себе нечувствительна к свету.
П. Бойсен-Йенсен установил, что если отрезать верхушку колеоптиля, а затем снова насадить ее так, чтобы между верхушкой и отрезанной частью поместилась бы прослойка из желатина или агара, то при освещении получатся такие же изгибы, как у нормальных колеоптилей. Следовательно, фототропическое раздражение передается через прослойку агара или желатина. Значит, верхушка проростка поставляет некое химическое вещество, и его перемещение определяет изгиб колеоптиля при одностороннем освещении.
Н. Г. Холодный, повторяя и модифицируя опыты Ч. Дарвина, предположил, что при геотропических изгибах важно наличие верхушки корня. В ней тоже образуется гормон, который перемещается от верхушки в нижележащую зону корня. Н. Г. Холодный и Ф. Вент дали объяснение изгибам проростков и корней. Они создали независимо друг от друга гормональную теорию тропизма и роста растений. Суть ее в следующем: под влиянием одностороннего освещения гормон смещается на затененную сторону проростка. Повышение его концентрации вызывает усиление роста, и проросток изгибается по направлению к свету.
Ф. Вент придумал связан оригинальный метод определения основного гормона роста - ауксина. Срезанная верхушка, положенная на агар, способна отдавать ему находящийся в ней гормон. Крошечный кубик из такого агара со стороной всего 2 миллиметра и расположенный на колеоптиле с отрезанной верхушкой только не прямо, а смещенный в сторону вызывает через час рост и искривление колеоптиля овса. По углу искривления судят о концентрации гормона. Чем больше угол, тем больше концентрация гормона
2. Ауксины. Строение, содержание, синтез, распределение в различных частях растений.Окислительный распад. Полярный транспорт.
Ауксины — это вещества индольной природы. Основным фитогормоном типа ауксина является b-индолилуксусная кислота (ИУК). Наиболее богаты ауксинами растущие части растительного организма. Образование ауксинов в большинстве случаев идет в меристематических тканях. Ауксины передвигаются из верхушки побега вниз к его основанию, а далее от основания корня к его окончанию. Таким образом, передвижение ауксинов полярно. Полярное передвижение ауксинов идет по проводящим пучкам со скоростью, значительно превышающей скорость обычной диффузии. Недостаток кислорода, торможение процесса дыхания с помощью различных ингибиторов приостанавливают передвижение ауксинов. Во взрослом дифференцированном растении при высокой концентрации гормона может наблюдаться и неполярное передвижение ауксинов вверх по растению с током воды по ксилеме. Ауксин, образующийся в кончике корня, может, по-видимому, передвигаться на короткие расстояния вверх, в зону растяжения. Основным источником для образования b-индолилуксусной кислоты (ИУК) является аминокислота триптофан. Синтез ИУК идёт как минимум в три стадии: декарбоксилирование, дезаминирование, окисление. ИУК может синтезироваться из индола и индолглицерофосфата. Именно в апикальной меристеме сосредоточен синтез ауксинов. Содержание ИУК зависит не только от скорости образования, но и от быстроты разрушения. Основным ферментом разрушения ИУК является ИУК-оксидаза (ОИУК). Наряду с ферментативным окислением ИУК большое значение имеет ее разрушение на свету (фотоокисление). Другим путем разрушения ИУК является декарбоксилирование. В клетках присутствует конъюгированный, т. е. связанный ауксина, который, как правило, неактивен. В клетках ауксин содержится в цитозоле и хлоропластах. Основными факторами, влияющими на содержание ауксина в растительных клетках являются следующие: триптофан-зависимый синтез ауксина, триптофан-независимый синтез ауксина, транспорт, окисление и декарбоксилирование, конъюгация. Образование ИУК зависит от снабжения растения азотом, обеспечения растения водой. Освещение уменьшает содержание ауксинов, а затемнение увеличивает. Под влиянием микроорганизмов содержание ауксинов у высших растений заметно возрастает.
3. Ауксины. Физиологическая активность и механизмы действия. Явление апикального доминирования. Практическое использование ауксинов в растениеводстве и биотехнологии.
Наиболее ярким проявлением физиологического действия ауксина является его влияние на рост клеток в фазе растяжения. ИУК стимулирует выход протонов в клеточную стенку и увеличивает ее растяжимость. Под влиянием ауксинов может измениться направление дифференциации клеток. Ауксины, так же как и другие фитогормоны, обусловливают взаимодействие отдельных органов растения (коррелятивный рост). Ауксин обуславливает явление апикального доминирования, проявляющегося в подавляющем влиянии верхушечной почки на рост пазушных. При всех физиологических проявлениях ауксины усиливают поступление воды и питательных веществ (аттрагирующее влияние). Ауксины влияют на распределение питательных веществ в растении. В отсутствие кислорода действие ауксина или не проявляется, или значительно ослабевает. ИУК увеличивает энергетическую эффективность дыхания растений. Под влиянием ИУК возрастает энергетический заряд клетки.ИУК активирует протонную помпу в плазмалемме, что приводит к закислению и разрыхлению клеточной стенки и тем самым способствует росту клеток растяжением. Комплекс ИУК с рецептором транспортируется в ядро и активирует синтез РНК, что в свою очередь приводит к усилению синтеза белков.
С участием ауксина связана регуляция двигательной активности растений, в частности тропизмы и настии. Повышение концентрации ауксина выше оптимальной вызывает торможение роста. Ауксин стимулирует синтез другого фитогормона, ингибирующего рост, этилена. Гормоны типа ауксина вызывают перераспределение питательных веществ в растении.
Ауксин стимулирует развитие бессемянных плодов. Использование, как гербицида. Если ауксинов слишком много, то начинается синтез гормона-антагониста этилена. Этилен угнетает рост побегов и корней в длину и вызывает листопад. При обработке естественными ауксинами (ИУК, ИПВК и др.) начинается их окисление и/или образование конъюгатов.
4. Гиббереллины. Открытие, строение, содержание, транспорт и распределение в различных частях и органах растений. Физиологическая активность и механизмы действия. Практическое применение.
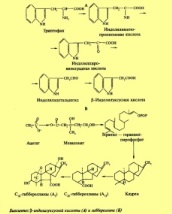 Открытие гормонов растений гиббереллинов связано с изучением болезни риса. Японские ученые показали, что эта болезнь вызывается выделением гриба Gibberella fujikuroi. Из выделений этого гриба было получено кристаллическое вещество — гиббереллин. Гиббереллины обладают высокой физиологической активностью и являются естественными фитогормонами. По химической структуре это производные дитерпенов — дитерпеноиды, состоящие из четырех изопреновых остатков. Растения на разных этапах онтогенеза могут различаться по набору гиббереллинов, активность которых может быть различной. Гиббереллины могут образовываться в разных, по преимуществу растущих частях растительного организма. Все же основное место синтеза гиббереллинов — это листья. Имеются данные, что гиббереллины образуются в пластидах. По-видимому, гиббереллины существуют в двух формах — свободной и связанной. Нередко наблюдаемое повышение содержания гиббереллинов связано с переходом их из связанной в свободную форму. В отличие от ауксинов гиббереллины передвигаются из листьев как вверх, так и вниз, как по ксилеме, так и по флоэме. Это пассивный процесс, не связанный с метаболизмом. Образование гиббереллина в хлоропластах идет путем превращения мевалоновой кислоты в геранилгераниол и далее через каурен в гибберелловую кислоту. Влияние факторов. Освещение увеличивает содержание гиббереллинов. При выращивании растений на красном свете в них содержится больше гиббереллинов по сравнению с выращиванием на синем свете. Улучшение питания растений азотом содержание гиббереллинов снижает.
Открытие гормонов растений гиббереллинов связано с изучением болезни риса. Японские ученые показали, что эта болезнь вызывается выделением гриба Gibberella fujikuroi. Из выделений этого гриба было получено кристаллическое вещество — гиббереллин. Гиббереллины обладают высокой физиологической активностью и являются естественными фитогормонами. По химической структуре это производные дитерпенов — дитерпеноиды, состоящие из четырех изопреновых остатков. Растения на разных этапах онтогенеза могут различаться по набору гиббереллинов, активность которых может быть различной. Гиббереллины могут образовываться в разных, по преимуществу растущих частях растительного организма. Все же основное место синтеза гиббереллинов — это листья. Имеются данные, что гиббереллины образуются в пластидах. По-видимому, гиббереллины существуют в двух формах — свободной и связанной. Нередко наблюдаемое повышение содержания гиббереллинов связано с переходом их из связанной в свободную форму. В отличие от ауксинов гиббереллины передвигаются из листьев как вверх, так и вниз, как по ксилеме, так и по флоэме. Это пассивный процесс, не связанный с метаболизмом. Образование гиббереллина в хлоропластах идет путем превращения мевалоновой кислоты в геранилгераниол и далее через каурен в гибберелловую кислоту. Влияние факторов. Освещение увеличивает содержание гиббереллинов. При выращивании растений на красном свете в них содержится больше гиббереллинов по сравнению с выращиванием на синем свете. Улучшение питания растений азотом содержание гиббереллинов снижает.
Проявлением физиологического действия гиббереллина является способность резко усиливать рост стебля у карликовых форм растений. Генетическая карликовость вызвана изменениями на генном уровне (отсутствие гена который кодирует синтез гиббереллинов) и может быть связана с нарушениями в синтезе гиббереллинов. 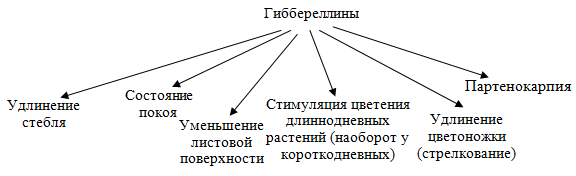 Партенокарпия-образование на растении плодов без оплодотворения
Партенокарпия-образование на растении плодов без оплодотворения
Гиббереллины заметно усиливают вытягивание стебля и у многих нормальных растений. Существует определенная зависимость между скоростью роста стебля растений и содержанием гиббереллинов. Увеличение роста стебля происходит за счет усиления деления клеток и их растяжения. Влияние гиббереллинов на растяжение связано с образованием белка клеточной стенки экстенсина и повышением активности ферментов. Гиббереллины накапливаются в почках при выходе из покоящегося состояния. При выходе семян из покоящегося состояния в них накапливаются гиббереллины. При действии гиббереллина возрастает общая масса растительного организма, он способствует не перераспределению питательных веществ, а общему их накоплению. Показано, что гиббереллин усиливает процесс фотосинтетического фосфорилирования, в первую очередь нециклического, и, как следствие, основных продуктов этого процесса — АТФ и НАДФН. Одновременно наблюдается снижение содержания хлорофилла. Следовательно, под влиянием гиббереллина повышается интенсивность использования единицы хлорофилла, возрастает ассимиляционное число. В темноте гиббереллин воздействует лишь на растяжение клеток, не вызывая возрастания интенсивности их деления.
5. Цитокинины. Природные и синтетические. Открытие, строение, содержание, места синтеза, транспорт и распределение в растениях. Физиологическая активность и механизмы действия. Взаимодействие с другими гормонами.
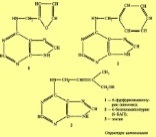 Учение о цитокининах начинается с открытия полуискусственного продукта – кинетина, который образуется из ДНК при кислотном гидролизе. Природный цитокинин был идентифицирован как зеатин и получен он из неспелых зерновок кукурузы.
Учение о цитокининах начинается с открытия полуискусственного продукта – кинетина, который образуется из ДНК при кислотном гидролизе. Природный цитокинин был идентифицирован как зеатин и получен он из неспелых зерновок кукурузы.
Открытие цитокининов связано с обширными исследованиями по выращиванию каллуса, образовавшегося из изолированной ткани сердцевины стебля табака на питательной среде. Было показано, что клетки каллуса в стерильной культуре через определенный промежуток времени прекращают деление. Однако при добавлении к питательной среде производных ДНК, получающихся после ее автоклавирования, деление клеток возобновляется. Все известные цитокинины — это производные пуриновых азотистых оснований, а именно аденина, в котором аминогруппа в шестом положении замещена различными радикалами.
Соединения цитокининового типа обнаруживаются в растениях не только в свободном состоянии, но и в составе некоторых тРНК. Богаты цитокининами клетки апикальных побегов и меристем корня. Цитокинины образуются главным образом в корнях и пассивно в виде зеатинрибозида передвигаются в надземные органы по ксилеме. Вместе с тем имеются данные об образовании цитокининов в семенах (зрелые зародыши) и развивающихся плодах.
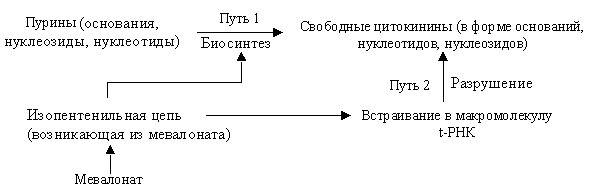 Ключевой фермент синтеза цитокининов — изопентенилтрансфераза и назван ipt-ген. Она катализирует синтез зеатина и рибозидзеатина из изопентенилпирофосфата. Изопентениловый остаток может образовываться из мевалоновой кислоты. Содержание цитокининов определяется скоростью их синтеза и разложения. Распад цитокининов регулируется ферментом цитокининоксидазой. Имеются сведения, что улучшение питания растений азотом усиливает образование цитокининов.
Ключевой фермент синтеза цитокининов — изопентенилтрансфераза и назван ipt-ген. Она катализирует синтез зеатина и рибозидзеатина из изопентенилпирофосфата. Изопентениловый остаток может образовываться из мевалоновой кислоты. Содержание цитокининов определяется скоростью их синтеза и разложения. Распад цитокининов регулируется ферментом цитокининоксидазой. Имеются сведения, что улучшение питания растений азотом усиливает образование цитокининов.
Основное место синтеза цитокининов – корни; однако в последнее время получены данные о том, что синтез цитокининов может происходить и в семенах (семена гороха).
Из корней цитокинины пассивно транспортируются в наземные органы по ксилеме.








