 2015-04-06
2015-04-06 8068
8068Дробление это ряд быстрых последовательных митотических делений зиготы. В промежутке между делениями клетки не растут, в результате зародыш становится многоклеточным, а по размеру не превышает зиготу.
Биологический смысл. Выравнивание ядерно-плазменного отношения. Достижене многоклеточности. Закладка основных осей симметрии (у большинства).
Зигота – Морула (от лат. Ягода тутового дерева) – Бластула (с полостью)
Клетки Бластулы – Бластомеры. Полости разделения – борозды дробления
Митоз:интерфаза=1:3 (или более)
Интерфаза = пресинтетический(предшественники ДНК, АТФ) + синтетический(синтез ДНК) +постсинтетический(синтез белков, АТФ) периоды
Во время дробления – есть запасы АТФ, питательных веществ, белков, предшественников ДНК. Во время дробления выпадают пресинтетический и постсинтетический периоды. Синтетический период каждого цикла по времени совпадает с телофазой предыдущего деления.
Первые дробления – синхронные. Постепенно, по мере истощения запаса пит.веществ нарастает асинхронность. Это связано с удлинением интерфазы (начинаются процессы синтеза). Момент когда начинаются собственные синтетические процессы – смена функции материнского генома зародышевым. Гибель – если сперматозоид несет летальные гены. Чем больше запас пит.веществ, тем позднее включается зародышевый геном.
|
|
|
Характер дробления зависит от количества желтка и его распределения в цитоплазме яйца
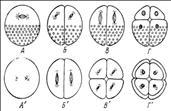
Правило Гертвига-Сакса: клеточное ядро стремится занять центр чистой, свободной от желтка цитоплазмы. Ось веретена дробления обычно совпадает с направлением наибольшей протяженности свободной от желтка цитоплазмы.
Гомолецитальный – равномерное распределение, представители ланцетник, иглокожие.
Телолецитальные – смещенное к краю, представители птицы.
Центролецитальные – в центре, представители членистоногие.
Типы дробления в зависимости от количества и распределения желтка в цитоплазме.
| дробление | |||
| голобластический (полное) | меробластический (частичное) | ||
| равномерная (бластомеры одинаковой величины -гомолецитальные) | неравномерные (не одинаковой величины - телолецитальные) | дискоидальное (дробится 1 диск -телолецитальные) | поверхностное (центролецитальное) |
| целобластула | амфибластула | дискобластула | перибластула |
Голобластическое дробление по расположению бластомеров:
· Радиальное – синхронно делятся
· Спиральное – после каждого деления, 2 дочерние клетки поворачиваются на 45°. Стерробластула. Черви и моллюски
· Билатеральное – имеется одна плоскость симметрии. Типично для аскариды.
· Анархическое – бластомеры слабо связаны между собой, сначала образуют цепочки или бесформенную массу. Кишечнополостные.
|
|
|
· Чередующиеся – асинхронность со второго дробления. Контактирующих частей больше. Бластоциста. Млекопитающие.
В результате дробления зародыш становится многоклеточным и готов к дальнейшей дифференциации.
1.Отдел Голосемянные.Отличия от высших споровых растений.Класс Хвойные, строение, размножение, значение. Голосеменные – древняя группа растений, появившаяся еще в девоне.Отдел современных голосеменных насчитывает более 700 видов. Современные голосеменные представлены преимущественно деревьями, значительно реже — кустарниками и очень редко —лианами; травянистых растений среди них нет. Листья голосеменных у большинства видов они игловидные (хвоя) или чешуевидные; Подавляющее большинство голосеменных — вечнозеленые, одно- или двудомные растения с хорошо развитыми стеблем и корневой системой, образованной главным и боковым корнями. Расселяются они семенами, которые формируются из семязачатков. Семязачатки голые (отсюда название отдела), расположены на мегаспорофиллах или на семенных чешуях, собранных в женские шишки. В цикле развития голосеменных наблюдается последовательная смена двух поколений — спорофита и гаметофита с господством спорофита. Гаметофиты сильно редуцированы, причем мужские гаметофиты голо- и покрытосеменных растений не имеют антеридиев, чем резко отличаются от всех разноспоровых бессеменных растений. Отличия от высших споровых растений. 1) появление семязачатка.2) Размножение семенами.3) Абсолютная разноспоровость. 4) Полная потеря связи с водной средой для оплототворения. 5)Замаскированност чередования поколений(гаметофита и спорофита).6) ВСР заросток самостоятельный (талломный), а у голосеменных м. и ж. находятся внутри семязачатка.7) Длительн. вторичный рост стебля, с обр. массивным древесиной с годичными кольцами.8) Все голосемянные имеют древесный образ жизни. Класс Хвойные, строение, размножение, значение. Все хвойные — вечнозеленые, реже листопадные (например, лиственница) деревья или кустарники с игольчатыми или чешуевидными {например, у кипариса) листьями. Игловидные листья (хвоя) плотные, кожистые и жесткие, покрыты толстым слоем кутикулы. Хвоя имеет ксероморфную структуру. Все эти особенности строения листьев обеспечивают хорошее приспособление хвойных к произрастанию как в засушливых, так и в холодных местообитаниях. Органы размножения хвойных растений — стробилы. Это видоизменённые укороченные побеги, на которых находятся специализированные листья — спорофиллы, формирующие спорангии — спорообразующие органы. У хвойных растений стробилы однополые, то есть содержат либо мужские, либо женские генеративные органы. Мужские стробилы называются микростробилами, чаще всего они растут компактными собраниями (иногда употребляется слово «соцветия», хотя с точки зрения ботаники это некорректно, поскольку стробилы не являются цветами) и только у некоторых примитивных хвойных — поодиночке. Отдельнорастущий микростробил или компактное собрание часто называют мужской шишкой. Обычно группы мужских шишек располагаются в пазухах листьев, реже — на верхушках боковых побегов. Мужские спорофиллы (микроспорофиллы) сильно редуцированны и имеют чешуевидную или щитковидную форму. На каждом микроспорофилле образуется 2—15 микроспорангиев, внутри микроспорангия находится спорогенная ткань, окруженная тапетумом — слоем клеток, доставляющих к ней питательные вещества. Из спорогенной ткани образуются микроспоры. Обычно число микроспор очень велико. Из каждой микроспоры развивается мужской гаметофит, у одних растений это происходит ещё внутри микроспорангия, у других — уже после попадания на женский стробил. Женские стробилы называются мегастробилами, чаще они растут поодиночке, реже — компактными собраниями (например, у тиса). Отдельнорастущий мегастробил или компактное собрание часто называют женской шишкой. Мегастробилы состоят из центральной оси и кроющих чешуй на ней, в пазухе каждой из кроющих чешуй расположена семенная чешуя, на которой, в свою очередь, сидят семязачатки, каждый из которых представляет собой нуцеллус — мегаспорангий, окруженный защитным веществом — интегументом. В каждом нуцеллусе находится 3—4 мегаспоры, однако только одна из них развивается в женский гаметофит — эндосперм. Большинство хвойных — однодомные., то есть на одном растении находятся как мужские, так и женские шишки. Двудомные хвойные встречаются реже. Хвойные растения — самые распространенные и имеющие наибольшее экономическое значение представители голосеменных, однако, они представляют собой лишь одну из четырёх подгрупп растений.
|
|
|
2.Современные представления о структуре и свойствах мембраны возбудимых клеток. Потенциал покоя, или мембранный потенциал и метод его регистрации. Природа потенциала покоя. Основным свойством живых клеток является раздражимость, т. е. их способность реагировать изменением обмена веществ в ответ на действие раздражителей. Возбудимость — свойство клеток отвечать на раздражение возбуждением.К возбудимым относят нервные, мышечные и некоторые секреторные клетки. Одним из важных свойств живых клеток является их электрическая возбудимость, т.е. способность возбуждаться в ответ на действие электрического тока. Возбуждение — ответ ткани на ее раздражение, проявляющийся в специфической для нее функции (проведение возбуждения нервной тканью, сокращение мышцы, секреция железы) и неспецифических реакциях (генерация потенциала действия, метаболические изменения). Строение и функции клеточных мембран. 1.Барьерная функция выражается в том, что мембрана при помощи соответствующих механизмов участвует в создании концентрационных градиентов, препятствуя свободной диффузии. При этом мембрана принимает участие в механизмах электрогенеза. К ним относятся механизмы создания потенциала покоя, генерация потенциала действия, механизмы распространения биоэлектрических импульсов по однородной и неоднородной возбудимым структурам. 2.Регуляторная функция клеточной мембраны заключается в тонкой регуляции внутриклеточного содержимого и внутриклеточных реакций за счет рецепции внеклеточных биологически активных веществ, что приводит к изменению активности ферментных систем мембраны и запуску механизмов вторичных «месенджеров» («посредников»).3.Преобразование внешних стимулов неэлектрической природы в электрические сигналы (в рецепторах). 4.Высвобождение нейромедиаторов в синаптических окончаниях. Потенциал покоя – это разность между электрическими потенциалами внутри и вне клетки в состоянии покоя.
Величина ПП варьируется в пределах 30-90 мВ.
ПП составляет основу возбуждения и переработки информации нервной клеткой, обеспечивает регуляцию деятельности внутренних органов и опорно-двигательного аппарата посредством запуска процессов возбуждения и сокращения в мышце. мембранный потенциал возбудимой клетки в невозбужденном состоянии.Он представляет собой разность электрических потенциалов, имеющихся на внутренней и наружной сторонах мембраны и составляет у теплокровных от -55 до -100 мВ. Известно, что в невозбужденном состоянии клеточная мембрана высокопроницаема для ионов калия и малопроницаема для ионов натрия. Установлено, что имеется разность концентраций ионов калия вне и внутри клетки, причем внутри клетки ионов калия примерно в 20—50 раз больше, чем вне клетки.
|
|
|
3.Типы взаимодействия аллельных генов (доминирование, кодоминирование, множественный аллелизм). Аллельным называют взаимодействие между аллельными генами. Доминирование - простейшая форма взаимодействия - была открыта еще Менделем. При доминировании доминантный ген полностью подавляет проявление рецессивного гена. Например, при скрещивании растений гороха с круглыми АА и морщинистыми аа образуются только круглые семена в первом поколении, во втором происходит расщепление в соотношении 3 круглых на 1 морщинистый.
Рецессивный аллель гена может проявиться в результате мутации. Измененный ген либо не кодирует белок, либо кодирует белок меньшей активности. В данном примере рецессивный генотип не влияет на жизнеспособность, но если белок необходим для жизни данного организма, то мутантный ген является летальным. Доминантные летальные аллели существуют в генофонде популяций многих видов. Рецессивные летальные гены не проявляются у гетерозиготного организма. Подсчитано, что человек гетерозиготен по 30 летальным рецессивным генам. Это объясняет тот факт, почему между потомками от браков среди близких родственников наблюдают высокую частоту летальных исходов. Кодоминированием называют проявление в гетерозиготном состоянии признаков, обусловленных действием обоих аллелей, т. е. каждый из аллельных генов кодирует определенный белок, а в гетерозиготном состоянии проявляются оба.
По типу кодоминирования у человека наследуются группы крови. Так, аллели Ia и Ibкодоминанты. У гетерозигот Ia Ib присутствуют оба генных продукта. Аллели Ia Ib доминируют над рецессивной аллелью I. Поэтому у человека система АВО определяет 4 группы крови: A- Ia Ib или IaI0, B-IbIb или IbI0, AB- Ia Ib но группа 0 - i0i0. Множественный аллелизм –это наличие нескольких аллелей одного гена. В популяции оказываются не два аллельных гена, а несколько.Множественный аллелизм для генов, контролирующих системы несовместимости, выступает как фактор отбора, препятствующий образованию зигот и организмов определенных зигот. Примером множественного аллелизма является серия множественных аллелей s1, s2, s3, обеспечивающих самостерильность многих растений. Двенадцать различных состояний одного локуса у дрозофилы, обусловливающих разнообразие окраски глаз (w — белые, we — эозиновые, wa — абрикосовые, wch — вишневые, wm — пятнистые и т. д.); серия множественных аллелей окраски шерсти у кроликов («сплошная», гималайская, альбинос и т. д.); аллели IA, Iв, I°, определяющие группы крови у человека, и т. д. Серия множественных аллелей — результат мутирования одного гена.

