 2015-05-12
2015-05-12 1203
1203Наконец, ФУС отвергает представление о поступательном ходе возбуждения по дуге рефлекса. В соответствии с этим представлением реализацию поведения обеспечивает активация последовательно включающихся в реакцию структур мозга. Сначала сенсорных структур, обрабатывающих сенсорную информацию, затем эф-фекторных структур, которые формируют возбуждение, активирующее железы, мышцы и т. д. Однако результаты многочисленных экспериментов показали, что при реализации поведенческого акта наблюдается не последовательное включение афферентных и эфферентных структур, а синхронная активация нейронов, расположенных в самых разных областях мозга. Паттерн активаций нейронов в этих структурах оказывается общим, имеет общемозговой характер. Компоненты этого паттерна — последовательные фазы активаций -- соответствуют последовательности развертывания описанных выше системных механизмов.
Таким образом, вовлечение нейронов разных областей мозга в системные процессы происходит синхронно. Эти процессы — общемозговые и не могут быть локализованы в какой-либо области мозга. В различных областях мозга в поведении протекают не локальные афферентные или эфферентные, а одни и те же общемозго-
вые системные процессы организации активности нейронов в систему, коте является не сенсорной или моторной, а функциональной. Активность нейронов этих областей отражает не обработку сенсорной информации или процессы регуляции движений, а вовлечение нейро в определенные фазы организации (афферентный синтез и принятие решения) и реализации системы. Активность лю структуры одновременно соответствует определенным свойствам среды, так характеру двигательной активности.
Единый паттерн активаций и синхронность вовлечения нейронов разных областей мозга в общемозговые системные процессы не означают эквипотенциальное™ (равнозначности) мозговых структур: вклад этих структур в обеспечение поведения зависит от специфики проекции на них индивидуального опыта (см. ниже).
Поведение
как континуум результатов
До сих пор мы с дидактическими целями, а также следуя традиции исходного варианта ФУС, использовали понятие пускового стимула. Однако ясно, что использование этого понятия в рамках парадигмы активности ведет к эклектике. Кажущаяся его необходимость отпадает при рассмотрении поведенческого акта не изолированно, а как элемента поведенческого континуума, последовательности поведенческих актов, совершаемых индивидом на протяжении его жизни. При этом оказывается, что следующий акт в континууме реализуется после достижения и оценки результата предыдущего акта. Эта оценка — необходимый компонент процессов организации следующего акта, которые могут быть рассмотрены как трансформационные или процессы перехода от одного акта к другому. Места для стимула в континууме нет (см. рис. 2.3). С изменениями среды, которые традиционно рассматриваются как стимул для данного акта, информационно связано на самом деле предыдущее поведение, в рамках которого эти изменения ожидались, предвиделись в составе модели будущего результата — цели.
2.2. Основы системной психофизиологии
А что же с неожиданными изменениями? К каким модификациям на уровне последовательности поведенческих актов может привести изменение среды, которое не предвиделось в рамках предшествующего ему поведения и, следовательно, не является результатом последнего? Оно либо не изменит запланированной последовательности актов континуума (и в этом смысле проигнорируется»), либо прервет ее, обус-онив формирование разных в зависимости! конкретной ситуации видов поведения: юр прерванного акта, формирование.'кого поведения для достижения требуе-результатов, в том числе и новых, 1. И опять это поведение будет направ-1 в будущее и его организация явится 'формационным эквивалентом будуще-о бытия.
Таким образом, поведение может быть актеризовано как континуум результа-(П.К. Анохин), а поведенческий акт мотрен как отрезок поведенческого пнуума от одного результата до другого Швырков).
Г 2.3. Системная детерминация • 'ивности нейрона
цигма реактивности: нейрон, индивид, отвечает на стимул
Как мы уже отмечали, с позиций пара-
чы реактивности поведение индивида
-тавляет собой реакцию на стимул.
„нове реакции лежит проведение воз-
!ия по рефлекторной дуге: от рецеп-
через центральные структуры к
лнительным органам. Нейрон при
жазывается элементом, входящим в
жторную дугу, а его функция — обес-
;ем проведения возбуждения. Тогда
енно логично рассмотреть детер-
лю активности этого элемента сле-
•1 образом: «...ответ на стимул,
; вовавший на некоторую часть ее
ой клетки. -- Ю.А.) поверхности,
распространяться дальше по клет-
йствовать как стимул на другие
ie клетки...» (Ф. Бринк, 1960, с. 93).
ательно, в рамках парадигмы реак-
:и рассмотрение нейрона вполне
методологически последовательно: нейрон, как и организм, реагирует на стимулы. В качестве стимула выступает импульса-ция, которую нейрон получает от других клеток как реакцию — следующая за си-наптическим притоком импульсация данного нейрона (рис. 2.4).
К сожалению, такая методологическая последовательность отсутствовала в рамках парадигмы активности. Как правило, анализ «нейронных механизмов» целенаправленного поведения приводил авторов к тому, что мы назвали выше «уровневой эклектикой»: представлению о том, что индивид осуществляет целенаправленное поведение, а его отдельный элемент -нейрон — реагирует на приходящее к нему возбуждение — стимул. Важнейшей задачей стало устранение подобной эклектики.
Критика понимания нейрона как проводника возбуждения
Подход к нейрону как к проводнику возбуждения встречал возражения уже давно, например, со стороны Дж.Э. Когхилла, который, однако, не мог в отсутствие целостной и последовательной теории, вписывающейся в парадигму активности, дать решение, адекватное сформулированной задаче. Его нейрон реагирует «на окружающую среду так же, как живой организм». Решающий шаг в направлении решения этой задачи был сделан П.К. Анохиным, который в своей последней работе подверг аргументированной критике общепринятую, как он ее назвал, «прове-денческую концепцию» нейрона и предложил вместо нее системную концепцию интегративной деятельности нейрона.
Вне зависимости от конкретных, усложняющихся с развитием науки представлений о функционировании нейрона в традиционном рассмотрении центральной оставалась идея об электрической сумма-ции потенциалов на мембране нейрона. В соответствии с ней предполагалось, что возбуждающие и тормозные постсинап-тические потенциалы, возникающие на мембране постсинаптического («получающего») нейрона под действием пресинап-тической импульсации за счет изменения
2. психоФи:
 |
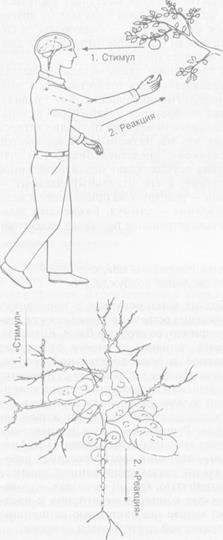 |
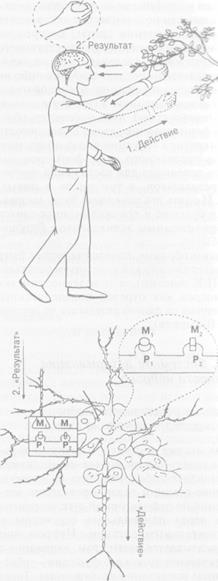
Рис. 2.4. Индивид и нейрон в парадигмах активности и реактивности
Цифрами на схеме обозначена последовательность событий. В соответствии с парадигмой реактивности
за стимулом (1) следует реакция (2) — поведенческая у человека, импульсная у нейрона. В последнем
случае в качестве стимула рассматривается импульсация нейрона, аксон которого (параллельный стрелке
с надписью «Стимул») контактирует с дендритом реагирующего нейрона, окруженного глиальными
клетками и соприкасающегося с кровеносным сосудом, который расположен над словом «Реакция».
Реакция — это импульсные разряды реагирующего нейрона. В соответствии с парадигмой активности
действие (1) (поведенческое у человека, импульсация у нейрона) завершается достижением результата
и его оценкой (2). Пунктиром обозначена модель будущего результата: для человека — контакт
с объектом-целью, для нейрона — получение соответствующих метаболитов (М1 — от контактирующего
нейрона, М2 — от соседней глиальной клетки), соединяющихся с рецепторами нейрона (P1f P2)
2.2. Основы системной психофизиологии
ионных градиентов, суммируясь, действуют на генераторный пункт нейрона, про-:>цирующий распространяющиеся потенциалы действия — импульсы.
П.К.Анохин назвал парадоксальным
еренос с нервного волокна на нейрон
.ставления о проведении возбуждения
лавной деятельности последнего. Если
ча состоит лишь в том, чтобы передать
хждение от одного нейрона к другому,
: неясно, для чего между входным и вы-
ым импульсами «вставлены» сложные
межуточные этапы: выделение медиа-
, его воздействие на субсинаптическую
'рану и химические превращения в ней.
жели для того, чтобы, начав с элект-
ского потенциала терминали, сформи-
гь в конце концов тот же спайковый
жал, весьма сходный по своим
ическим параметрам с потенциалом,
мшим по аксонной терминали?» -
:ивал он [Анохин, 1975, с. 368].
-"он - организм,
'. -ающий необходимые метаболиты
:зоей «микросреды»
еупомянутые этапы приобретают том случае, если принять, что про-: обеспечивающий переход от пре- к штическим образованиям, продол-и непрерывную цепь химических ов внутри нейрона и — главное -
межклеточные контакты служат метаболическими субстратами меж-
ктирующими клеточными образо-[. Переход от «проведенческой кон-
к рассмотрению нейрона как ор-. получающего необходимые ему
1 гы из окружающей «микросреды», •м шагом, который предопределил
)щую разработку проблемы в на-
и ее системного решения.
~\ia активности: нейрон, чцивид, изменяя соотношение ~осредой», удовлетворяет свои Юности»
"ходимость дальнейшей разработки -пась тем, что в рамках концепции деятельности нейрона по-
следовательность событий в принципе оставалась такой же, как и в парадигме реактивности. В обоих случаях процесс начинался приходом возбуждения к нейрону и заканчивался генерацией этим нейроном потенциала действия. Разница, которую подчеркивал П.К. Анохин, состояла в том, какими процессами заполнялся интервал между действием медиатора на субсинаптическую мембрану нейрона и генерацией потенциала: химическими преобразованиями внутри нейрона в первом случае и электрической суммацией — во втором.
Устранение эклектики и приведение представления о детерминации активности нейрона в соответствие требованиям системной парадигмы было достигнуто отказом от рассмотрения активности нейронов как реакции на синаптический приток и принятием положения о том, что нейрон, как и любая живая клетка, реализует генетическую программу, нуждаясь в метаболитах, поступающих к нему от других клеток (В.Б. Швырков). В связи с этим последовательность событий в деятельности нейрона становится аналогичной той, которая характеризует активный целенаправленный организм, а его импульсация -- аналогичной действию индивида (см. рис. 2.4).
Активность нейрона, как и поведение организма, является не реакцией, а средством изменения соотношения со средой, «действием», которое обусловливает устранение несоответствия между «потребностями» и микросредой, в частности, за счет изменения синаптического притока. Это изменение, если оно соответствует текущим метаболическим «потребностям» нейрона, приводит к достижению им «результата» и прекращению активности. Предполагается, что несоответствие между «потребностями», определяемыми генетически, и реально поступающими метаболитами может иметь место как при генетически обусловленных изменениях метаболизма клетки, так и при изменении притока метаболитов от других клеток. Таким образом, нейрон не «кодирующий элемент», «проводник» или «сумматор», а организм в организме, обеспечивающий свои «потребности» за счет метаболитов, поступающих от других элементов.
2. ПСИХОФИЗИОЛОГИЯ
Направленность в будущее и обусловленность метаболическими «потребностями» активности нейрона как необходимые компоненты ее системного понимания
Следует подчеркнуть, что для последовательно системного понимания детерминации активности нейрона существенны оба компонента: признание направленности активности нейрона в будущее и ее обусловленности метаболическими «потребностями» нейрона. То, что только первого из них недостаточно, видно на примере интересной концепции гедонистического нейрона, разработанной [KJopf, 1982]. Утверждая, что целенаправленный мозг состоит из целенаправленных нейронов, автор этой концепции отвечает на вопрос «В чем нейроны нуждаются и как они это получают?», исходя из следующей логики. Аристотель рассматривал получение удовольствия как главную цель поведения. Следовательно, организм гедонистичен. Нейрон есть организм. Следовательно, нейрон гедонистичен. «Удовольствие» для нейрона — возбуждение, а «неудовольствие» — торможение. Активация нейрона — «действие», обеспечивающее получение им возбуждения. Нейрон является гетероста-том — системой, направленной "на максимизацию «удовольствия», т. е. возбуждения.
Таким образом, отсутствие второго из двух необходимых компонентов позволяет предположить, что у нейрона имеются довольно странные и экзотически аргументированные «потребности». Особенно если принять во внимание популярную концепцию «токсического перевозбуждения» (excitotoxic), в рамках которой сильное возбуждение нейронов рассматривается как причина их гибели.
В то же время наиболее часто в концепциях авторов отсутствует первый компонент. А это при анализе нейронного обеспечения поведения ведет к рассмотрению сложных метаболических превращений внутри нейрона главным образом как фактора, обеспечивающего проведение возбуждения и пластичность (модификацию проведения при разных видах научения). При этом сложнейшие механизмы изменения
«белкового фенотипа» оказываются направленными, например, на изменение чувствительности постсинаптической мембраны к пресинаптической импульсации.
«Потребности» нейрона
Охарактеризуем очень кратко некоторые существенные «потребности» нейрона. Они определяются необходимостью синтеза новых молекул, в том числе белков, расходуемых в процессе жизнедеятельности («типичная» белковая молекула разрушается в среднем через два дня после того, как она была синтезирована) или обеспечивающих структурные перестройки нейрона при обучении. Для этого в том случае, если в клетке нет соответствующей информационной РНК, направляющей синтез белка в цитоплазме, экспрессируются (становятся активными, «выраженными») гены, среди которых выделяют гены «домашнего хозяйства» (универсальные «потребности» клеток), гены «роскоши» (специфические «потребности» клетки) или «ранние» и «поздние» гены, экспрессируемые на последовательных стадиях формирования памяти, и т. д. Как предполагается, именно усложнение процессов регуляции экспрессии генов (а не их числа) определяет эволюционное усложнение живых систем.
Различие в экспрессии (а не потеря или приобретение генов) определяет различие специализации клеток организма. Особенно велики эти различия для клеток мозга, в которых экспрессируются десятки тысяч уникальных для мозга генов. Считается, что метаболическая гетерогенность нейронов, обусловленная генетически и зависящая от условий индивидуального развития, т. е. являющаяся результатом взаимодействия фило- и онтогенетической памяти, лежит в основе разнообразия функциональной специализации нейронов, определяет специфику их участия в обеспечении поведения.
Объединение нейронов в систему
как способ обеспечения метаболических
«потребностей»
Нейрон может обеспечить «потребности» своего метаболизма только объединяясь с другими элементами организма в функ-
2.2. Основы системной психофизиологии
циональную систему. Их взаимосодействие, совместная активность обеспечивает достижение результата, нового соотношения целостного индивида и среды. «Изнутри», на уровне отдельных нейронов, достижение результата выступает как удовлетворение метаболических «потребностей» нейронов и прекращает их активность.
Такой подход к пониманию активности нейронов заставил применить для ее анализа вместо традиционных «постстимуль-ных» гистограмм, выявляющих закономерные изменения активности нейрона после предъявления стимула, «предрезультат-ные» гистограммы, которые позволяют обнаружить нейроны, импульсная активность которых закономерно увеличивается при реализации поведения, направленного на получение данного результата, и прекращается при его достижении.
Важность системного понимания детерминации активности нейрона для психологии
Итак, поскольку системная психофизиология отвергает парадигму реактивности, основывая свои положения на представлении об опережающем отражении, о направленной в будущее активности не только индивида, но и отдельных нейронов, постольку она обеспечивает для психологии, оперирующей понятиями активности и целенаправленности, возможность избавиться от эклектических представлений, часто появляющихся при использовании материала нейронаук.
2.2.4. Субъективность отражения
Активность как субъективное отражение
Рассмотрение соотношения индивида и среды с позиций ФУС уже давно привело к заключению о том, что поведенческий континуум целиком занят процессами организации и реализации функциональных систем (см. рис. 2.3); специального временного интервала для процессов обработки сенсорной информации не обнаружива-
ется. Представление о том, что индивид не реагирует на стимулы, кодируя и декодируя информацию об их свойствах, а реализует активность, направленную в будущее, т. е. опережающее отражение, связанное с формированием внутренней субъективной модели будущего события ~ результата, с необходимостью требует признания отражения субъективным.
На определенном этапе казалось, что коротколатентные активации некоторых нейронов могут быть сопоставлены с кодированием физических параметров стимула для последующего сличения с имеющейся в памяти моделью. Однако скоро стало ясно, что даже самые ранние активации нейронов в поведенческом акте — не кодирование, а уже результат сличения с субъективными моделями, сформированными в рамках предыдущего акта континуума.
Сказанное находится в соответствии с положением о «пристрастности» отражения среды, о зависимости последнего от целей поведения и имеющегося у индивида опыта. Это свойство психического отражения обозначается как субъективность и предполагает несводимость описания отражения к языку сенсорных модальностей, выражающих в «сенсорном коде» физические параметры объектов.
Опережающее отражение вместо обработки информации для построения «образов-картинок»
В четкой форме опережающий характер отражения представлен в когнитивной психологии концепцией У. Найссера, который считает, что образы не «картинки в голове», появляющиеся после действия сенсорных стимулов, а «предвосхищения будущего». Автор подчеркивает, что предвосхищение не обязательно является реалистическим.
Действительно, мы можем рассмотреть наш опыт как состоящий из актов-гипотез, включающих параметры планируемых результатов, отношения между ними, пути их достижения и т. д. Гипотезы тестируются во внутреннем и внешнем плане. И хотя можно полагать, что отбор из ряда «проб-
2. ПСИХОФИЗИОЛОГИЯ
ных» актов «удачного», попадающего в видовую память, определяется соответствием гипотезы реальным свойствам и закономерностям среды, тем не менее возможность достижения конкретным индивидом в том или ином поведенческом акте требуемого соотношения организма и среды, т. е. результата, не означает, что данная гипотеза целиком базируется на упомянутых свойствах и закономерностях. В экологической психологии убедительные аргументы против того, что среда состоит из стимулов и отображается как «картинка», рассматриваемая гомунку-люсом, приведены Дж. Гибсоном. Им разработана стройная теория, которая, как справедливо замечает А.Д. Логвиненко (см. [Гибсон, 1988]), в руководствах либо игнорируется, либо искажается до неузнаваемости в связи с невозможностью ее ассимилировать, оставаясь в рамках традиционной парадигмы. И это не удивительно, так как принципиальными положениями этой теории является отрицание не только схемы «стимул — реакция», но и самого понятия «стимул». Автор отвергает также идею о необходимости обработки и передачи сенсорной информации -ее некому принимать. Ниже мы еще вернемся к теории Дж. Гибсона.
Физические характеристики среды и целенаправленное поведение
Более 30 лет назад Дж. Леттвин с соавторами, изучив связь активности нейронов сетчатки лягушки с поведением, сформулировали в очень яркой форме свое представление о том, что выделяет организм в среде: «лягушки интересуются жуками и мухами, в то время как границы и углы интересуют только ученых». Еще раньше в гештальтпсихологии были обоснованы положения о том, что среда должна определяться не физически, а психобиологи-чески и что целостное восприятие не составляется из отдельных элементарных «кусков». «Куски», физические характеристики, в соответствии с которыми ранжируются стимулы и связь с которыми устанавливается при анализе активности нейронов или отчетов испытуемых, появ-
ляются в результате специального поведения, направленного на выделение этих характеристик: классификация, сравнение объектов, например, в науке, искусстве и т. п. Следует согласиться с Дж. Гибсоном в том, что объект не складывается из качеств, но мы можем выделить их, если это надо для целей эксперимента. На что же мы «дробим» среду, что выделяем в ней, если не упомянутые физические характеристики?
Теория эффордансов Дж. Гибсона
С позиций парадигмы активности с давних пор представлялось очевидным, что из среды активно «отбирается» индивидом то, что может быть использовано для достижения цели, причем, как считал Ж. Икс-кюлль, число объектов, которые может различить индивид, равно числу функций, которые он может реализовать. Анализ среды как обеспечивающей активность индивида в ней, дан в теории affordance Дж. Гибсона. Неологизм «affordance» (эф-форданс) — существительное, образованное Гибсоном от глагола «afford» — предоставлять. Эффордансы — это то, что окружающий мир предоставляет, разрешает индивиду, чем он его обеспечивает для совершения поведения. Эффордансы нельзя предъявить индивиду, так как они не являются стимулами. Можно лишь обеспечить их наличие. Автор считает, что индивид соотносится не с миром, описываемым в физических терминах, а с экологическим миром. Он понимает экологические нишу вида как набор эффордансов. Понятие «эффорданс» подразумевает взаимодополнительность мира и индивида. Гибсон отмечает, что понимает под ним «нечто, относящееся одновременно и к окружающему миру, и к животному таким образом, который не передается ни одним из существующих терминов» [Гибсон, 1988, с. 188].
«Дробление» среды индивидом
как отражение истории их соотношения
Как мы уже знаем, основным понятием в ФУС является результат, под которым понимается соотношение организма и среды и который, следовательно, так же, как
2.2. Основы системной психофизиологии
эффорданс, относится одновременно к окружающему миру и к индивиду. Однако в отличие от эффорданса результат, как и валентность у К. Левина, включает субъективный компонент, от которого отказывается Гибсон, постулируя независимость эффордансов от потребностей и опыта наблюдателя. Поэтому для ответа на вопрос о том, как дробит среду индивид, как она представлена в его субъективном мире, мы должны дополнить экологический мир субъективным компонентом, т. е. подчеркнуть аспект использования эффордансов индивидом.
При этом оказывается, что среда дробится тем или иным образом в соответствии с опытом совершения индивидом тех или иных поведенческих актов на протяжении его индивидуального развития. Индивид отражает не внешний мир как таковой, а историю своих соотношений с миром. Описание среды индивидом основано на оценках его соотношения с объектами-целями поведенческих актов, т. е. на оценках результатов. Образно говоря, можно рассматривать жизнь индивида как «ассимиляцию» экологического мира, превращающую для индивида экологический мир в мир результатов. Продолжая данную логику, можно заключить, что среда представлена для индивида результатами реализованных актов.
В этой части излагаемая здесь система представлений довольно близко примыкает к концепции У. Матурана [1996]. Он считает, что мнение об организме, имеющем входы и выходы, и о нервной системе, кодирующей информацию об окружающей среде, не выдерживает критики. Понятие информации Матурана относит к «степени неуверенности наблюдателя в своем поведении в области определенных им самим альтернатив» [там же, с. 137]. Автор справедливо утверждает, что состояния активности репрезентируют отношения (между организмом и средой), а не являются описанием окружающей среды. Это описание может быть дано исключительно в терминах, содержащихся в «когнитивной области наблюдателя», в терминах поведения организма. Тогда «знать — значит уметь вести себя адекватно в ситуациях, связанных с индивидуальными актами» [там же, с. 135].
Субъективность отражения и зависимость системной организации активности центральных и периферических нейронов от цели поведения
Убедительные примеры, демонстрирующие проявление субъективности отражения в организации активности мозга, можно получить при анализе зависимости от цели поведения активности нейронов «сенсорных» структур, которую принято считать детерминированной модально-специфической стимуляцией.

