 2015-05-12
2015-05-12 6472
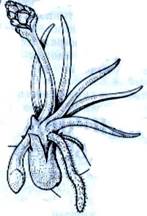
6472Палеонтологический материал свидетельствует, что плаунообразные являются самой древней группой среди современных высших растений. Достоверные остатки их известны с верхнего силура палеозойской эры. Массовое расселение плаунообразных произошло в девонском периоде. Чередование поколений в жизненном цикле оказалось весьма полезным, поскольку позволило часть жизни проводить во влажных условиях (гаметофит), а другую часть – в более сухих. Сухопутный образ жизни спорофита способствовал развитию надземной части растения. Наибольшего развития споровые достигли в каменноугольном периоде. Вместе с хвощеобразными и папоротникообразными, плаунообразные составляли основу растительного покрова Земли. Наряду с травянистыми формами, некоторые плаунообразные были мощными деревьями – лепидодендроны (Lepidodendron) и другие (рис. 4). Большинство их достигало в высоту 10–35 м (отдельные экземпляры до 45 м). Более низкорослые – сигиллярии (Sigillaria).
Условия обитания растений каменноугольного периода сильно отличались от современных. В световом и тепловом отношении они были более однотипными. Суша занимала относительно небольшую поверхность и омывалась безбрежной теплой акваторией. Она многократно пересекалась широкими полноводными реками; господствовали низменности, покрытые мелководными морями, водоемами и заболоченными пространствами. На поверхность суши эффективно действовало также внутреннее тепло Земли. До предела насыщенная парами туманная атмосфера непроницаемым слоем окутывала Землю, защищала планету от прямых солнечных лучей и потери тепла через излучение. Все это способствовало выравниванию климата земной поверхности. Климат нынешних умеренных областей Европы и Северной Америки был равномерно влажный тропический и субтропический.
Растения развивались круглый год. На мелководьях, заболоченных и илистых, пропитанных водой почвах, формировался густой растительный покров. Однако в таких условиях формируется поверхностная корневая система и высокие растения были подвержены ветровалу. Подмытые водой, особенно во время разливов, деревья падали, покрывались наносами. Огромные скопления их подвергались разложению без доступа воздуха и превращались в многометровые толщи каменного угля. Остатки высокорослых плаунообразных и хвощеобразных образовали основные запасы каменного угля Европы, Северной Америки, частично Азии. К концу карбона большинство древовидных форм споровых растений вымирает, уступив место новым, более прогрессивным группам растений, в первую очередь голосеменным.
Современные плаунообразные представляют собой угасающую группу высших растений, включающую около 1 000 видов, относимых к 10 родам. Их участие в сложении растительного покрова незначительное.
Ископаемые и современные плаунообразные обычно типично листостебельные растения и представлены различными жизненными формами спорофита. Наиболее примитивные и древние плаунообразные – первично травянистые формы (астероксилон (Asteroxylon), схизоподиум (Schizopodium), протолепидодендрон (Protolepidodendron) и
др.). В девонском периоде возникают вторично древесные формы, достигшие, как отмечено, максимального развития в карбоне. В мезозое их сменили невысокие (2–3 м) древовидные формы – плевромейя (Pleuromeia). В настоящее время сохранились только травянистые формы, как первично (плаун (Lycopodium), баранец (Huperzia), филлоглоссум (Phylloglossum), селягинелла (Selaginella) и др.), так и вторично травянистые – полушник (Isoёtes), стилитес (Stylites); обычно вечнозеленые, многолетние (рис. 4).
Подземные органы некоторых ископаемых и современных форм имеют вид типичного корневища (отдельные плауны). Часто образуется своеобразный орган, несущий спирально расположенные корни – ризофор (от греч. rhiza – корень и phoros – несущий), или корненосец (лепидодендрон, сигиллярия, плевромейя, полушник). Нередко их называют стигмариями (от греч. stigma – клеймо), поскольку на них хорошо заметны округлые следы (рубчики) отходивших ранее корней. Иногда формируются надземные корненосцы – селагинелла. Редко подземная часть представлена клубнем – филлоглоссум. Для корней плаунообразных характерен самый древний примитивный тип ветвления – дихотомический. Формируются корни первого, второго и последующего порядков. Дихотомическое ветвление возникло у растений, обитавших на рыхлой или насыщенной водой почве. Корни не проникали далеко в глубь почвы. Современные растения этого отдела сохранили тип ветвления корней предков. Они лишены корневых волосков. Корни плаунообразных – придаточные или адвентивные. Как и у всех высших споровых растений главный корень отсутствует. Корневая система их – первично гоморизная.
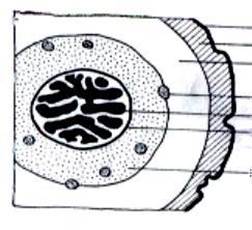
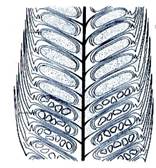
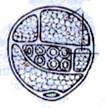
Рис. 4. Плаунообразные (Lycopodiophyta): 1 – Астероксилон (Asteroxylon); 2 – филлоид баррагванатии (Barragwanathia) с проводящим пучком; 3 – побег дрепанофикуса (Drepanophycus); 4 – схема поперечного среза стебля плауна булавовидного (Lycopodium clavatum), (вк – внутренняя кора, к – ксилема, лс – листовые следы, нк – наружная кора, ск – средняя кора, э – эпидерма, эн – эндодерма); 5 – филлоглоссум (Phylloglossum), (б – боковой вырост клубня, к – корни, кл – клубень); 6 – протолепидодендрон (Protolepidodendron); 7 – лепидодендрон (Lepidodendron); 8 – сигиллярия (Sigillaria), (с – стигмарии, ст – стробилы); 9 – фрагмент гетероспорового стробила лепидостробуса, или флемингитиса (Lepidostrobus – Flemingites), (мг – мегаспорангии, мк – микроспорангии); 10 – плевромейя (Pleuromeia); 11 – «семя» миадесмии (Miadesmia), (м – мегаспорофилл); 12а–12в – селягинелла (Selaginella), (12а – мужской гаметофит: кс – клетки стенки антеридия, пк – проталлиальная клетка, ск – сперматогенные клетки; 12б – женский гаметофит с зародышем: з – зародыш, ом – оболочка мегаспоры, р – ризоиды; 12в – зародыш: г – гаустория, п – подвесок, пк – первичный корень, пч – почечка); 13 – натхорстиана (Nathorstiana); 14 – нижняя часть спорофилла полушника озерного (Isoёtes lacustris), (с – спорангий, я – язычок); 15 – стилитес (Stylites).

э
 нк
нк
ск
2 лс
3 к
4 эн
1 вк
 |


ст
 7 8 с
7 8 с
б
к

 6 кл м
6 кл м
 кс
кс
 мк
мк
ск
 11 12а пк
11 12а пк

 мг з
мг з
10 пч п пк
12 б ом
 я 12в г
я 12в г
 с
с
14 13 15
Корни плаунов формируются крайне своеобразно – у развивающегося из зиготы зародыша зачатки побега и корня возникают одновременно в результате дихотомирования точки роста. В процессе развития растения новые корни образуют из меристемы апекса побега. Заложение и развитие корня и побега как двух равноценных органов подтверждается сходством анатомического строения базальной части корня и стебля. При заглублении корня в почву его анатомическое строение изменяется в соответствии с функциями и спецификой среды обитания. Эндогенно закладываются корни у селягинелловых на ризофорах или на стебле. Формирование корневых систем плаунообразных показывает, что стеблевые придаточные корни в эволюции растительного мира возникли раньше главного. Они характерны для всех высших споровых растений.
Побеги современных плаунообразных прямостоячие, стелющиеся, полегающие, у эпифитов – свисающие. Они также ветвятся дихотомически (равно и неравно). Сочетание двух типов дихотомического ветвления наблюдается как у современных, так и у ископаемых древесных и травянистых форм (нередко на одном растении – дифазиаструм сплюснутый (Diphasiastrum complanatum)). Иногда неравная дихотомия приводит к формированию мощной прямой главной оси (псевдомоноподиальное ветвление астероксилона, плауна поникшего (Lycopodium cernuum)). В процессе эволюционного упрощения древесных и травянистых форм возникают слабо разветвленные (сигиллярия) и неразветвленные растения (плевромейя, натхорстиана (Nathorstiana), полушник, стилитес, филлоглоссум).
Проводящая система плаунообразных представлена различными типами стели. У вымерших травянистых форм – гаплостела и актиностела (астероксилон). Вымершие древесные растения характеризовались наличием сифоностели. Современные травянистые формы также характеризуются разнообразием типов стели. Для плаунов характерна актиностела и плектостела (проводящие ткани располагаются в виде лент, дающих анастомозы). Встречается также сифоностела (филлоглоссум), актиностела (полушник) и др. У древнейших представителей отдела ксилема в актиностеле была мезархной, у современных – она экзархная.
Многолетие побеги современных плаунообразных не нарастают в толщину (нет камбия), за исключением полушника. Вымершие древесные и некоторые травянистые формы характеризовались вторичным утолщением стеблей и ризофоров.
Водопроводящие элементы ксилемы представлены обычно трахеидами разных типов (кольчатые, спиральные, поровые, с разными типами боковой поровости), но и настоящими сосудами (некоторые селягинеллы). Проводящие элементы флоэмы – чаще длинные ситовидные клетки.
Проводящая система плаунообразных небольшая по размерам. Основную массу стебля (до 90% и больше) составляет мощная кора, которая нередко дифференцируется на несколько зон. Наиболее сложная по строению кора у ископаемых древесных форм (наряду с первичными тканями хорошо развиты вторичные ткани, образованные феллогеном, или пробковым камбием). Формировались «коревые деревья».
Плаунообразные характеризуются микрофиллией, что является их важнейшей особенностью. Листьям (филлоидам) свойственна анатомическая и морфологическая простота строения. По мнению многих исследователей, в процессе эволюционного развития они сформировались из поверхностных выростов осей – энациев. По размерам – сравнительно мелкие. Листовая пластинка цельная, с одной простой жилкой, реже на верхушке – дихотомически разветвленной (протолепидодендрон и др.). Основание часто несколько расширенное, иногда в виде подушечки. Она представлена группой рыхлых клеток, пронизанных системой межклетников (аэренхима), связанной аэренхимой стебля. Форма листа – чаще шиловидная, лентовидная, чешуевидная. Прорывы в стеле над листовыми следами отсутствуют. Листорасположение чаще очередное.
У вымерших форм класса Селягинеллоподобные (Selaginellopsida) на верхней поверхности расширенного основания листа формировались выросты в виде язычка, или лигулы для удаления излишков влаги. У большинства современных представителей родов Селагинелла, Полушник имеется его рудимент. У вегетативных листьев он быстро засыхает, сохраняется лишь у спорофиллов. По-видимому, у современных видов лигула не имеет биологического значения.
Энационное происхождение листьев плаунообразных подтверждается также особенностями их онтогенеза – лист развивается из одной поверхностной клетки стебля. Первые листья молодого растения не имеют проводящего пучка.
В жизненном цикле доминирует спорофит (см. рис. 1). По строению спор плаунообразные подразделяются на две группы: равно- и разноспоровые. У равноспоровых (астероксилон, баррагванатия (Barragwanathia), дрепанофикус (Drepanophycus), баранец, дифазиаструм (Diphasiastrun), ликоподиелла (Lycopodiella), плаун, филлоглоссум, протолепидодендрон и др.) все споры одинакового размера. У плаунов они мелкие – диаметром около 30 мкм. У разноспоровых форм (лепидодендрон, лепидокарпон (Lepidocarpon), плевромейя, сигиллярия, селягинелла, полушник, стилитес и др.) имеются крупные мегаспоры диаметром от 90 до 1 500 мкм и мелкие многочисленные микроспоры, диаметр которых в среднем в десять раз меньше (от 20 до 100 мкм). Количество мегаспор в мегаспорангиях намного меньше. У селягинеллы от 4 до 1.
Таким образом, разноспоровость впервые возникает у плаунообразных: на ранних этапах эволюции сосудистых растений – в среднем девоне (более 360 млн. лет назад).
Спороносные структуры плаунообразных также отличаются большим разнообразием. У наиболее примитивных (баррагванатия) спорангии располагались непосредственно на стебле вперемежку с вегетативными листьями. В дальнейшем спорангии располагались в пазухах листьев (дрепанофикус), на листьях, у их основания (протолепидодендрон, баранец). На более поздних этапах дифференцируются спорофиллы (спорангии формируются на их верхней стороне), которые чередовались с вегетативными листьями (трофофиллами) – образуются спороносные зоны побегов. Следующий этап – формирование специализированных спороносных колосков, или стробилов (плаун, дифазиаструм, ликоподиелла). Последние формы составляют подавляющее число видов. У разноспоровых форм на микро- и мегаспорофиллах соответственно развиваются микро- и мегаспорангии. Также формируются стробилы на концах побегов. Спорофиллы имеют небольшой вырост – язычок. Спорангии располагаются в пазухах спорофиллов (между язычком и осью стробила). Стробилы бывают смешанными (лепидодендрон, сигиллярия, селягинелла) или односпоровыми. Спорангии многих таксонов плаунообразных – на массивной короткой ножке.
Плаунообразные – эуспорангиатые растения (спорангий развивается из группы клеток). Стенка спорангия 2–3 слойная. Самый внутренний слой – тапетум почти полностью разрушается. Содержимое его клеток используется при развитии спор. Содержимое развивающегося спорангия – многоклеточная спорогенная ткань (археспорий), при митотическом делении клеток которой, образуются материнские клетки спор – спороциты. Вследствие редукционного деления из них формируются тетрады спор. Созревшие споры обычно округло-тетраэдрической формы с двумя оболочками. Наружная оболочка (экзоспорий) плаунов скульптурированная, имеет часто сетчатый рисунок (с утолщенными гранями). Внутренняя оболочка (эндоспорий) тонкая. В цитоплазме споры содержится до 35% масла. В сухую погоду зрелые споры высыпаются из спорангия через образовавшуюся в его стенке трещину. При увеличении влажности она закрывается, предохраняя споры от намокания и преждевременного прорастания. Некоторые каменноугольные плаунообразные имели споры с воздушными мешками.
При прорастании спор формируется половое поколение (гаметофит), представленное заростком. Заростки равноспоровых и разноспоровых форм резко различаются. Заростки равноспоровых форм обоеполые массивные, длиной 2–20 мм, морфологически различающиеся у разных видов. По образу жизни они бывают подземными, полуподземными и наземными. У видов с подземными заростками (например, плаун булавовидный) споры прорастают через несколько лет после образования. Они постепенно погружаются в почву на глубину 1–8 см. Заросток развивается в течение 6–15 лет, однако его размеры составляют 2–5 мм. Еще на ранних стадиях развития в его ткани внедряются гифы некоторых почвенных грибов и располагаются в 3–5 слоях под нижней эпидермой. Питание заростка микотрофное.
Полуподземные заростки имеют хлорофиллоносную паренхиму в участках над поверхностью почвы. Но им также свойственен симбиоз с грибами. Характерно одновременно микотрофное и фототрофное, т.е. совмещенный тип питания. Развиваются в течение 5-7 лет.
Надземные заростки ряда тропических видов с хорошо развитой хлоренхимой. Они являются автотрофами. Развиваются в течение одного вегетационного периода.
Заростки разноспоровых форм однополые и развиваются в течение нескольких дней или недель. Женские заростки более долговечные. Однако живут не более одного вегетационного периода. Развиваются за счет питательных веществ споры, и по достижении зрелости не выступают или слегка выступают за пределы оболочки споры. Мужские заростки редуцируются в большей степени, чем женские, вплоть до нескольких клеток.
Микро- и мегаспоры часто начинают прорастать внутри микро- и мегаспорангиев. Однако завершается полное развитие заростков обычно вне материнского организма, после отделения спорангиев.
В связи с этим следует отметить, что задержка развития гаметофита и нового молодого спорофита на материнском растении приводит к возникновению в мезозойской эре на основе разноспоровости семенных растений. Возникновение разноспоровости – важный шаг в эволюции высших растений.
На заростках развиваются органы полового процесса. Антеридии погружены в ткань заростка. Архегонии имеют хорошо развитое брюшко, погруженное в ткань заростка и короткую шейку, возвышающуюся над его поверхностью. В антеридиях формируются двух- или многожгутиковые сперматозоиды, в архегониях – яйцеклетки.
Оплодотворение совершается при наличии капельно-жидкой среды, что является лимитирующим фактором для местообитания. После оплодотворения, на женском заростке без периода покоя начинается развитие зиготы. Развивается небольшой подвесок, прикрепляющий зародыш в теле заростка. Зародыш в начале питается за счет заростка. По мере появления вегетативных органов (стебелька с листьями, придаточных корней) зародыш переходит к самостоятельной жизни. Заросток отмирает.
Вегетативное размножение у плаунов и селагинелл обычно происходит при распадении побегов на части и укоренении этих частей. У некоторых видов имеются специализированные органы вегетативного размножения: выводковые клубеньки, выводковые луковички или почки, столоновидные побеги. В результате разрастания стелющихся побегов плаунов, особь может занять достаточно большую территорию, формируя клон. У некоторых плауновых наблюдается вегетативное размножение гаметофита.
Классификация плаунообразных связана с большими трудностями из-за наличия переходных форм среди вымерших растений. Отдел чаще подразделяют на два класса: равноспоровые Плауноподобные, или Ликоподиопсиды и преимущественно разноспоровые Полушникоподобные, или Изоэтопсиды (Селягинеллоподобные).








