 2015-06-10
2015-06-10 2860
2860(рис. 12-3)
Неконъюгированный билирубин в плазме прочно связан с альбумином. Только очень небольшая часть билирубина способна подвергаться диализу, однако под влиянием веществ, конкурирующих с билирубином за связывание с альбумином (например, жирных кислот или органических анионов), она может увеличиваться. Это имеет важное значение у новорождённых, у которых ряд лекарств (например, сульфаниламиды и салицилаты) может облегчать диффузию билирубина в головной мозг и таким образом способствовать развитию ядерной желтухи.
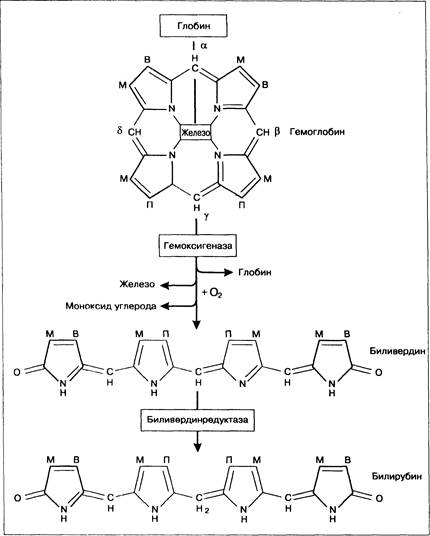
Рис. 12-2. Превращение гемоглобина в билирубин. М — метильная группа; В — винильная группа; П — пропионат.
Печенью выделяются многие органические анионы, в том числе жирные кислоты, жёлчные кислоты и другие компоненты жёлчи, не относящиеся к жёлчным кислотам, такие как билирубин (несмотря на его прочную связь с альбумином). Исследования показали, что билирубин отделяется от альбумина в синусоидах, диффундирует через слой воды на поверхности гепатоцита |55]. Высказанные ранее предположения о наличии рецепторов альбумина не подтвердились. Перенос билирубина через плазматическую мембрану внутрь гепатоцита осуществляется с помощью транспортных белков, например транспортного белка органических анионов [50], и/или по механизму «флип-флоп» [55]. Захват билирубина высокоэффективен благодаря его быстрому метаболизму в печени в реакции глюкуронидизации и выделению в жёлчь, а также вследствие наличия в цитозоле связывающих белков, таких как лигандины (глутатион-8-трансфераза).
Неконъюгированный билирубин представляет собой неполярное (жирорастворимое) вещество. В реакции конъюгации он превращается в полярное (водорастворимое вещество) и может благодаря этому выделяться в жёлчь. Эта реакция протекает с помощью микросомального фермента уридиндифосфатглюкуронилтрансферазы (УДФГТ), превращающего неконъюгированный билирубин в конъюгированный моно- и диглюкуронид билирубина. УДФГТ является одной из нескольких изоформ фермента, обеспечивающих конъюгацию эндогенных метаболитов, гормонов и нейротрансмиттеров.
Ген УДФГТ билирубина находится на 2-й паре хромосом. Структура гена сложная (рис. 12-4) [2, 54]. У всех изоформ УДФГТ постоянными компонентами являются экзоны 2—5 на 3'-конце ДНК гена. Для экспрессии гена необходимо вовлечение одного из нескольких первых экзонов. Так, для образования изоферментов билирубин-УДФГТ1*1 и 1*2 необходимо вовлечение соответственно экзонов 1А и ID. Изофермент 1*1 участвует в конъюгации практически всего билирубина, а изофермент 1*2 почти или вовсе не участвует в этом [25]. Другие экзоны (IF и 1G) кодируют изоформы фенол-УДФГТ. Таким образом, выбор одной из последовательностей экзона 1 определяет субстратную специфичность и свойства ферментов.
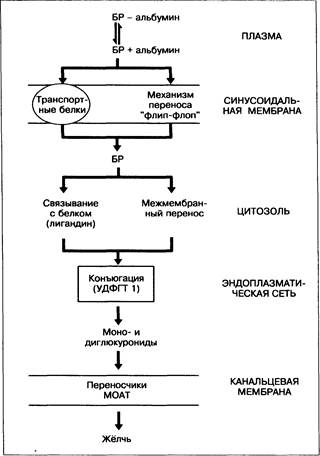
Рис. 12-3. Захват, метаболизм и секреция билирубина (БР). MOAT — мультиспецифичный транспортный белок для органических анионов.
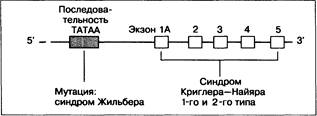
Рис. 12-4. Строение гена УДФГТ 1*1, содержащего 5 экзонов и промоторный участок (последовательность ТАТАА). Всего может быть 10 экзонов (показан только 1). Экзон 1А (называемый также экзоном 1*1) участвует в экспрессии УДФГТ 1*1 (основного активного фермента), экзон ID (или 1*4) — в экспрессии УДФГТ 1*2.
Дальнейшая экспрессия УДФГТ 1*1 зависит также от промоторного участка на 5'-конце, связанного с каждым из первых экзонов |6|. Промоторный участок содержит последовательность ТАТАА.
Детали строения гена важны для понимания патогенеза неконъюгированной гипербилирубинемии (синдромы Жильбера и Криглера—Найяра; см. соответствующие разделы), когда в печени содержание ферментов, ответственных за конъюгацию, снижено или они отсутствуют.
Активность УДФГТ при печёночно-клеточной желтухе поддерживается на достаточном уровне, а при холестазе даже увеличивается. У новорождённых активность УДФГТ низкая.
У человека в жёлчи билирубин представлен в основном д и глюкуронидом. Превращение билирубина в моноглюкуронид, а также в диглюкуронид происходит в одной и той же микросомальной системе глюкуронилтрансферазы [37]. При перегрузке билирубином, например при гемолизе, образуется преимущественно моноглюкуронид, а при уменьшении поступления билирубина или при индукции фермента возрастает содержание диглюкуронида.
Наиболее важное значение имеет конъюгация с глюкуроновой кислотой, однако небольшое количество билирубина конъюгируется с сульфатами, ксилозой и глюкозой; при холестазе эти процессы усиливаются [II].
В поздних стадиях холестатической или печёночно-клеточной желтухи, несмотря на высокое содержание в плазме, билирубин в моче не выявляется. Очевидно, причиной этого является образование билирубина типа III, моноконъюгированного, который ковалентно связан с альбумином [54]. Он не фильтруется в клубочках и, следовательно, не появляется в моче. Это снижает практическую значимость проб, применяемых для определения содержания билирубина в моче.
Экскреция билирубина в канальцы происходит с помощью семейства АТФ-зависимых мультиспецифичных транспортных белков для органических анионов [27]. Скорость транспорта билирубина из плазмы в жёлчь определяется этапом экскреции глюкуронида билирубина.
Жёлчные кислоты переносятся в жёлчь с помощью другого транспортного белка. Наличие разных механизмов транспорта билирубина и жёлчных кислот можно проиллюстрировать на примере синдрома Дубина—Джонсона, при котором нарушается экскреция конъюгированного билирубина, но сохраняется нормальная экскреция жёлчных кислот. Большая часть конъюгированного билирубина в жёлчи находится в смешанных мицеллах, содержащих холестерин, фосфолипиды и жёлчные кислоты. Значение аппарата Гольджи и микрофиламентов цитоскелета гепатоцитов для внутриклеточного транспорта конъюгированного билирубина пока не установлено.
Диглюкуронид билирубина, находящийся в жёлчи, водорастворим (полярная молекула), поэтому в тонкой кишке не всасывается. В толстой кишке конъюгированный билирубин подвергается гидролизу b-глюкуронидазами бактерий с образованием уробилиногенов. При бактериальном холангите часть диглюкуронида билирубина гидролизуется уже в жёлчных путях с последующей преципитацией билирубина. Этот процесс может иметь важное значение для образования билирубиновых жёлчных камней.
Уробилиноген, имея неполярную молекулу, хорошо всасывается в тонкой кишке и в минимальном количестве — в толстой. Небольшое количество уробилиногена, которое в норме всасывается, вновь экскретируется печенью и почками {энтерогепатическая циркуляция). При нарушении функции гепатоцитов печёночная реэкскреция уробилиногена нарушается и увеличивается почечная экскреция. Данный механизм объясняет уробилиногенурию при алкогольной болезни печени, при лихорадке, сердечной недостаточности, а также на ранних стадиях вирусного гепатита.








