2015-06-26
2015-06-26 5244
5244Химическое строение и свойства. В 20-е гг. прошлого столетия Г. Эванс сумел излечить бесплодие у содержащихся на синтетической диете крыс, добавляя им в корм листья салата. Активное соединение, способствующее развитию эмбриона, было выделено также из масел зародышей пшеницы и других семян. Оно получило название “токоферол” (от греч. tokos – потомство, phero – несу).
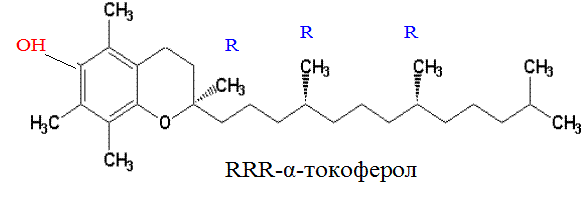
В 1938 г. токоферол, или витамин Е, был синтезирован химическим путем. Однако между природным и синтетическим витамином Е существуют различия, что отражено в их классификации. Натуральные формы токоферола обозначаются как RRR-a-токоферолы (R обозначает конфигурацию метильной группы) и имеют единственный стереоизомер; синтетические формы называются all-rac- a -токоферолы и являются рацемической смесью из 8 стереоизомеров, из которых 7 не найдено в природе. Основной биологически активной формой является RRR-α-токоферол:
Свободная ОН-группа обуславливает свойства витамина Е как антиоксиданта.
Кроме токоферолов к группе витамина Е относятся α, β, γ и δ-токотриенолы – аналоги соответствующих токоферолов, которые отличаются от последних наличием двойных связей в боковой цепи. Токоферолы – прозрачные, светло-желтые, вязкие масла, хорошо растворимые в большинстве органических растворителей. Медленно окисляются на воздухе, разрушаются под действием УФ-лучей.
Метаболизм. Витамин Е поступает в желудочно-кишечный тракт в составе масел, гидролиз которых липазой и эстеразой приводит к высвобождению витамина. Затем он всасывается и в составе хиломикронов поступает в лимфатическую систему и кровяное русло. В печени витамин связывается с токоферолсвязывающими белками, причем наибольшим сродством обладает RRR-α-токоферол. Другие токоферолы выделяются из печени с желчными кислотами. Эти белки “экспортируют” витамин в кровь в составе ЛПОНП. В плазме крови происходит обмен токоферолом между ЛПОНП и другими липопротеинами крови. Обмен между фракциями липопротеинов (особенно между ЛПНП и ЛПВП) и эритроцитами обеспечивает равновесие токоферола в крови.
Витамин поступает в экстрапеченочные ткани в составе ЛННП, которые захватываются соответствующими рецепторами. Кроме такого рецепторно-опосредованного механизма имеется и другой,– ферментативно-опосредованный, зависящий от активности липопротеинлипазы: фермент высвобождает токоферол из хиломикронов и ЛПОНП, после чего витамин поступает в ткани путем пассивной диффузии (рис.3.3). Благодаря пассивной диффузии через клеточную мембрану концентрация RRR-α-токоферола увеличивается во всех тканях организма, особенно в мозге. Структурная организация фосфолипидов клеточных мембранах способна “узнавать” хиральную форму RRR-α-токоферола, благодаря чему витамин задерживается в мембране, где и выполняет свою функцию (синтетические токоферолы в составе мембраны обеспечивают ее меньшую защиту от оксидативного стресса).
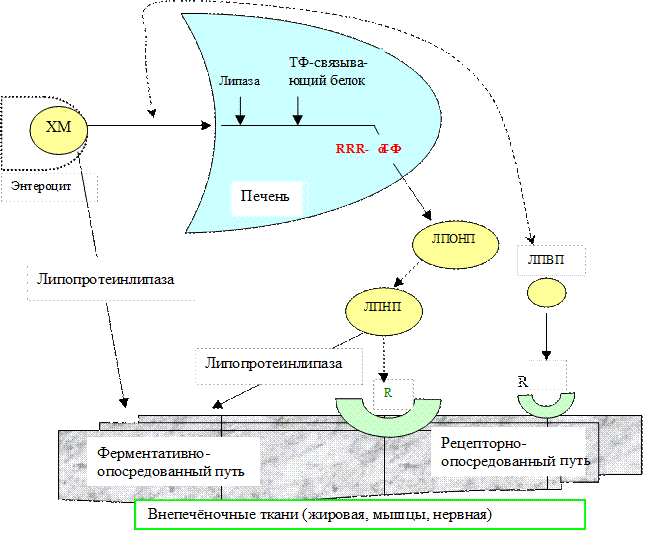
Рис.3.3. Транспорт витамина Е
Не всосавшиеся в кишечнике токоферолы выводятся с калом. Продукты метаболизма витамина – токофериновая кислота и ее водорастворимые глюкурониды – выводятся с мочой.
Биохимические функции. Биохимические функции токоферола многообразны.
1. Витамин Е является универсальным протектором клеточных мембран от окислительного повреждения. Он занимает такое положение в мембране, которое препятствует контакту кислорода с ненасыщенными липидами (образование гидрофобных комплексов). Это защищает биомембраны от их перекисной деструкции. Антиоксидантные свойства токоферола обусловлены также способностью подвижного гидроксила хроманового ядра его молекулы непосредственно взаимодействовать со свободными радикалами кислорода (О2˙, НО˙, НО2˙), свободными радикалами ненасыщенных жирных кислот (RO˙, RO2˙) и пероксидами жирных кислот. Мембраностабилизирующее действие витамина проявляется и в его свойстве предохранять от окисления SH-группы мембранных белков. Его антиоксидантное действие заключается также в способности защищать от окисления двойные связи в молекулах каротина и витамина А. Витамин Е (совместно с аскорбиновой кислотой),способствует включению селена в состав активного центра глутатионпероксидазы, тем самым он активизирует ферментативную антиоксидантную защиту(глутатионпероксидаза обезвреживает гидропероксиды липидов).
 | |||
 | |||
/ \ ООi
\ / / / \
 / \ _║¯\ + ОН-ТФ \ / + •O-ТФ
/ \ _║¯\ + ОН-ТФ \ / + •O-ТФ
 \ / / / / \
\ / / / / \
/ \‗/‾ \ \ /
\ / / \‗
/ \ \_
\ / \‗
/ \ \
\ / /
/ \ \
/
Пероксидный радикал ненасы- Ненасыщенная жир- радикал
сыщенной жирной кислоты Токоферол ная кислота токоферола
2. Токоферол является не только антиоксидантом, но и антигипоксантом, что объясняется его способностью стабилизировать митохондриальную мембрану и экономить потребление кислорода клетками. Следует отметить, что из всех клеточных органелл митохондрии наиболее чувствительны к повреждению, так как в них содержится больше всего легко окисляющихся ненасыщенных липидов. Вследствие мембраностабилизирующего эффекта витамина Е в митохондриях увеличивается сопряженность окислительного фосфорилирования, образование АТФ и креатинфосфата. Важно также отметить, что витамин контролирует биосинтез убихинона – компонента дыхательной цепи и главного антиоксиданта митохондрий.
3. Токоферол контролирует синтез нуклеиновых кислот (на уровне транскрипции), а также гема, микросомных цитохромов и других гем-содержащих белков.
4. Витамин Е обладает способностью угнетать активность фосфолипазы А2 лизосом, разрушающей фосфолипиды мембран. Повреждение мембран лизосом приводит к выходу в цитозоль протеолитических ферментов, которые и повреждают клетку.
5. Витамин Е является эффективным иммуномодулятором, способствующим укреплению иммунозащитных сил организма.
Гиповитаминоз. Недостаточность токоферола – весьма распространенное явление, особенно у людей, проживающих на загрязненных радионуклидами территориях, а также подвергающихся воздействию химических токсикантов. Глубокий гиповитаминоз встречается редко – преимущественно у недоношенных детей (проявляется гемолитической анемией).
При Е-витаминной недостаточности наблюдается частичный гемолиз эритроцитов, в них снижается активность ферментов антиоксидантной защиты. Повышение проницаемости мембран всех клеток и субклеточных структур, накопление в них продуктов ПОЛ – главное проявление гиповитаминоза. Именно этим обстоятельством объясняется разнообразие симптомов недостаточности токоферола. – от мышечной дистрофии и бесплодия вплоть до некроза печени и размягчения участков мозга, особенно мозжечка. Увеличение активности выходящих из поврежденных тканей ферментов в сыворотке крови (креатинфосфокиназы, аланинаминотрансферазы и др.) и увеличение содержания в ней продуктов ПОЛ наблюдается уже на ранних стадиях Е-гиповитаминоза.
Дефицит витамина Е в организме сопровождается снижением содержания иммуноглобулинов E. После его введения нормализуется численность Т- и В-лимфоцитов в периферической крови и восстанавливается функциональная активность Т-клеток.
Описаны врожденные нарушения обмена витамина Е, проявляющиеся анемией и мышечной дистрофией.
Оценка обеспеченности организма токоферолом. Основным методом является определение концентрации витамина Е в крови. Однако при гипер-β-липопротеинемиях (в том числе и возрастных) содержание токоферола в крови не отражает уровень обеспеченности им тканей организма (ЛПНП – одна из транспортных форм витамина).
Наиболее доступны клинической лаборатории функциональные тесты, позволяющие оценить глубину Е-витаминной недостаточности. Основными являются тесты на перокидную или гемолитическую устойчивость эритроцитов, определение суточной экскреции креатина (мышцы при Е-гиповитаминозе плохо «удерживают» креатин) и содержание продуктов ПОЛ в крови и эритроцитах.
Гипервитаминоз. Витамин не токсичен при значительных (10-20-кратных к суточной потребности) и длительных превышениях его дозировки, что обусловлено ограничением способности специфических токоферолсвязывающих белков печени включать витамин в состав ЛПОНП. Его избыток выводится из организма с желчью. В некоторых случаях длительный прием мегадоз токоферола (более 1г/сут) может привести к гипертриглицеридемии и повышению кровяного давления.
Суточная потребность. Пищевые источники. Основной источник токоферола – растительные масла, однако только свежие, получаемые методом холодного прессования. Поскольку растительное масло обычно экстрагируют из семян при высокой температуре, затем подвергают очистке, дезодорированию и рафинированию, оно значительно обедневает токоферолом. Более того, избыток растительных масел в рационе усиливает недостаточность витамина Е в организме, так как он расходуется на интенсифицированный потреблением ненасыщенных жирных кислот масел процесс ПОЛ.
Лучшим пищевым источником витамина Е являются орехи, семечки, гречневая крупа, проросшие ростки пшеницы. Он содержится в листьях салата и капусты. Из продуктов животного происхождения более всего токоферолов в сливочном масле, сале, мясе, желтке яиц. В молоке этого витамина мало.
Суточная потребность – 10 мг. Однако токоферол быстро расходуется в организме, особенно в условиях стимулированного ПОЛ, что имеет место при многих заболеваниях. Окислительной деструкции витамина Е препятствует витамин С. Это обусловлено присутствием в молекуле последнего енольной группы - донора водорода, который способен «гасить» свободный радикал токоферола. Тем самым витамин С экономит фонд витамина Е. С целью усиления антиоксидантного эффекта токоферола его следует назначать с аскорбиновой кислотой.
Витамин Д (кальциферол), антирахитический.
Химическое строение и свойства. В 1936 г. А. Виндаусом из рыбьего жира был выделен препарат, излечивающий рахит. Он был назван витамином Д3, так как ранее А. Гессом и М. Вейнштоком из растительных масел был выделен эргостерин, получивший название витамин Д1. При воздействии на витамин Д1 УФ-лучей образовывалось излечивающее рахит соединение – витамин Д2, эргокальциферол (кальциферол означает несущий кальций). В растениях при УФ-облучении синтезируются и другие витамеры эргостерина (Д4 -7).
Наиболее важным из группы витаминов Д является витамин Д3 –холекальциферол. Холекальциферол образуется из 7-дегидрохолестерола в клетках кожи человека под влиянием УФ-лучей.
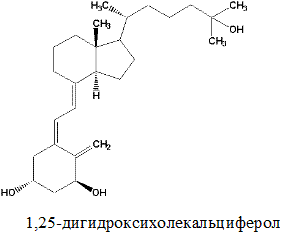
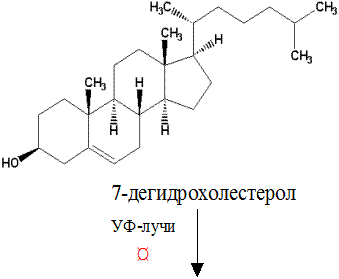
Витамин Д3 (холекальциферол)
Метаболизм. Витамины группы Д всасываются подобно витамину А (см. выше). В печени витамины подвергаются гидроксилированию микросомной системой оксигеназ по С-25 (из витамина Д3 образуется 25(ОН)-Д3, т.е. 25-гидроксихолекальциферол) и затем переносятся с помощью специфического транспортного белка с током крови в почки. В почках осуществляется вторая реакция гидроксилирования по С-1 с помощью митохондриальных оксигеназ (образуется 1,25(ОН)2-Д3, т.е. 1,25-дигидроксихолекальциферол, или кальцитриол). Эта реакция активируется паратиреоидным гормоном, секретируемым паращитовидной железой, когда уровень кальция в крови снижается. Если уровень кальция адекватен физиологической потребности организма, вторичное гидроксилирование происходит по С-24 (вместо С-1), при этом образуется неактивный метаболит 1,24(ОН)2-Д3.В реакциях гидроксилирования принимает участие витамин С.
Витамин Д3 накапливается в жировой ткани. Выводится главным образом с калом в неизмененном или окисленном виде, а также в виде конъюгатов.
Биохимические функции. Витамин Д3 можно рассматривать как про гормон, так как он превращается в 1,25(ОН)2-Д3, действующий аналогично стероидным гормонам. Так, проникая в клетки-мишени, он связывается с белковыми рецепторами, которые мигрируют в ядро клетки. В энтероцитах этот гормон-рецепторный комплекс стимулирует транскрипцию иРНК, несущую информацию на синтез белка-переносчика ионов кальция. Вероятно, витамин отвечает также за синтез Са2+-АТФ-азы в разных клетках.
В кишечнике всасывание кальция осуществляется как путем облегченной диффузии (с участием кальцийсвязывающего белка), так и путем активного транспорта (с помощью Са2+-АТФ-азы). Одновременно ускоряется и всасывание фосфора.
В костной ткани 1,25(ОН)2-Д3стимулирует процесс деминерализации (синергично с паратирином).
В почках активация витамином 1,25(ОН)2-Д3кальциевой АТФ-азы мембран почечных канальцев приводит к увеличению реабсорбции ионов кальция; возрастает и реабсорбция фосфатов.
Кальцитриол принимает участие в регуляции роста и дифференцировке клеток костного мозга. Он обладает антиоксидантным и антиканцерогенным действием.
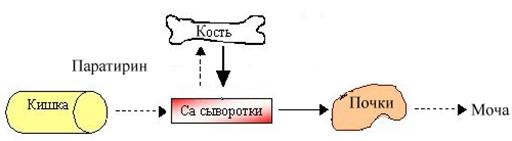
Гиповитаминоз. Недостаток витамина Д у детей приводит к заболеванию рахитом. Основные проявления этого заболевания сводятся к симптоматике недостаточности кальция. Прежде всего страдает остеогенез: отмечается деформация скелета конечностей (искривление их в результате размягчения – остеомаляции), черепа (позднее заращение родничков), грудной клетки (появление своеобразных «четок» на костно-хрящевой границе ребер), задерживается прорезывание зубов. Развивается гипотония мышц (увеличенный живот), возрастает нервно-мышечная возбудимость (у младенца выявляется симптом облысения затылочка из-за частого вращения головкой), возможно появление судорог. У взрослого недостаточность кальция в организме приводит к кариесу и остеомаляции (размягчение кости); у пожилых – к развитию остеопороза(снижение плотности костной ткани вследствие нарушения остеосинтеза). Разрушение неорганического матрикса объясняется усиленным «вымыванием» кальция из костной ткани и нарушением реабсорбции кальция в почечных канальцах при дефиците витамина Д:
На схеме показано угнетение (пунктирная стрелка) всасывания, снижение поступления кальция в кость и уменьшение экскреции кальция при недостатке витамина Д. Одновременно в ответ на гипокальциемию секретируется паратирин и увеличивается (сплошная стрелка) поступление кальция из кости в кровяное русло (вторичный гиперпаратиреоидизм).