 2015-06-28
2015-06-28 4860
4860Гендік мутацияларға сипаттама. Гендер мутациялары (нүктелі) барлық органикалық формаларда кездеседі. Жоғарыда айтылғанындай, олар жекелеген клеткаларда өтеді және жекелеген особьтарда (мутанттарда) сёкірмелі түрде байқалады.
Түрдің жабайы формаларына тән болатын гендер аллелін жабайы типі немесе қалыпты гендер деп, ал өзгергендерін —- мутантты гендер деп атайды. Олардың арасында принципті айырмашылық жоқ. Түрдің жабайы формаларына тән көптеген гендер де бір кезде мутантты гендер болған, одан соң қолайлы мутанттық аллельдер түр эволюциясының барысында сол түрге жататын особьтардың бәріне таралатындай концентрацияға жеткен.
Пайда болған кезде мутациялардың көпшілігі рецессивті.болып шығады. Түрдің тіршілігін сақтауда мүның өте зор ма-ңызы бар, өйткені жаңадан пайда болатын мутациялар көп ретте генотиптің тұтас системасын бүза отырып, зиянын тигізе-ді. Алайда олардың рецессивті сипатты болуы оларға түр особьтарына зиян келтірместен гетерозиготалы күйде сақталу-ға және келешекте гомозиготалы күйге өткен кезде көрінуге мүмкіндік береді.
Геннің жабайы типтен жаңа жағдайға мутациялануын — тура, ал мутанттықтан жабайы типке мутациялануын — кері мутация деп атайды. Ал кері мутацйялану процесінін, езін ген реверсиясы деп атайды. Бастапқы ген аралық сатысыз-ақ жаңа жағдайға және керісінші утацияланады. Тура мутацияның пайда болу жиілігі әр түрлі гендер үшін түрліше болады, орта есеппен алғанда 100 мың немесе 1 млн. генге бірден беске дейін тура мутация келеді, демек, мутациялар — өте сирек кездесетін құб.ылыс. Алайда әсімдіктер, жануарлар популяциялары мен адамда әр түрлі мутанттық гендердің кездесуінің жиынтық жиі-лігін ескергенде бұл цифр кенет артады. Мәселен, адамдардың әрбір гаметасында тіршілік қабілетін кемітетін 5—6-га дейін рецессивті мутанттық гендер болатынын арнайы есептеулер көрсетті (29-тарауды қара). Белгілі бір мутациялар әр түрлі уа-қытта пайда болуы мүмкін. Бұл гендердің бір бағытта әлденеше рет мутациялана алатынын білдіреді.
Кептік аллелизм. Осы уақытқа дейін материалды баяндауда біз гомологты хромосомалардың белгілі бір локусы екі аллель түрінде көрінуі мүмкін деген қағиданы басшылыққа алып кел-дік. Олар: А және а, В және е, С және с т. б. Іс жүзінде белгілі бір ген бірнеше күйге өзгере алады; кейде мүндай күй бірнеше ондаған және түпті жүздеген шамаға жетуі мүмкін. А гені а1, а2, аъ,...ап күйіне мутацияланады. Белгілі бір геннің бірқатар күйінжатқызады. көптік аллельдер сериясы деп, ал құбылыстың өзін көптік ал-леллизм деп атайды.
Көптік аллельдер сериясын зерттеу мұндай серияның кез келген аллелі жабайы типтегі аллельден немесе осы серияның кез келген басқа мүшесінен тікелей мутация жолымен пайда бо-ла алатынын, ал серияның әрбір мүшесінің, сірә өзіне тән мутациялану жиілігі болуы мүмкіндігін көрсетті.
Көптік аллельдер сериясы мүшелерінің түқым қуалауы Мендель заңдылықтарына бағынады (VII -таблица). Мүнда, оларға екі күй ғана белгілі болатъш гендерден езгеше, гетеро-зиготада көптік аллельдер сериясының әр түрлі екі мүшесінің үйлесуін компаунд деп атайды.
Көптік аллельдер сериялары мүйізді ірі қарадан, кроликтерден, тышқандардан, теңіз Біошқасынан, дрозифила шыбынынан, сондай-ақ жүгері, темекі, бүршақ және т. б. табылған. Адамда аллельдердің ІА, 1В, 1° сериялары белгілі, олар қан топтары бойынша мынадай полиморфизмді анықтайды: АВ тобы ІАІВ генотипіне сай келеді:
А» ІА ІА немесе ІА 1°»»
В» Ів Ів немесе Ів 1°»»
О» 1° 1°»»
Өсімдіктердің өздігінен стерыльділігін анықтайтын көптік аллельдер сериясы локусының болуы бірқат.ар жағдайларда айқас ұрықтануды қамтамасыз ететін механизм болып табыла-ды. Мәселен, темекіде, бедеде және басқа өсімдіктерде аналық аузында өздігінен стерильділік локусының аналық аузы генотипінде болатын аллельдерден өгеше аллелі бар тозаң ғана есетіні кәрсетілді.
Жануарлар, өсімдіктер мен микроорганизмдер арасында кептік аллелизмнің көп таралуы және оньщ адамда болуы бұлқүбылыс мутациялық өзгергіштік резервін арттыратындығына, осыған орай эволюцияда бейімделушілік маңызы бар екендігіне байланысты болуы мүмкін.
3. ХРОМОСОМАЛЫҚ МУТАЦИЯЛАР
Хромосомаішілік өзгерістер. Хромосомалардың қайта қүры-луын мутацияларға жатқызу қабылданған, өйткені клеткаларда олардың болуы әдетте осы клеткалар қасиеттерінің неме-се олардаы пайда болатын организмдердің өзгеруімен байла-нысты болады.
Хромосомаішілік езгерістерге хромосома бөліктерінің же-тіспеушілігін (дефишенсия және делеция); хромосоманың бел-гілі бір учаскелерінің екі еселенуін, немесе дәлірек айтқанда, кебеюін (дупликация); хромосоманың жекелеген учаскелерінің 180°-қа аударылуы салдарынан хромосомадағы гендердің сы-зықтық орналасуының өзгеруін хромосомаішілік өзгерістерге
Хромосома бөліктерінің жетіспеушілігі ұзындықтары әр
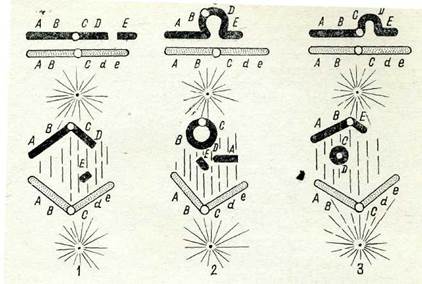
65-сурет. Хромосомалар жетіспеушілігінщ түрлері:
1 — үшынан жетіспеушілігі; 2 — ұштарынан жетіспеушіліктері; 3 — хромосомаішілік жетіспёушілік,
түрлі хромосома учаскелерін және оның түрлі бөліктерін қамтуы мүмкін. Егер хромосома иіндерінің бірінің соңы жойылатындай болып үзілсе (65-сурет, 1), онда осы иін қысқарады. Егер онда центромералар жоқ болса, бөлініп қалған үзінді ондағы гендерімен бірге ядроның таяудағы бөлінуі кезінде жойылады. Мүндай жетіспеушілікті терминальдық немесе соңы (концевыми) деп атайды.
Кейде хромосоманың бірден екі иіні үзіледі, осының салдарынан оның екі үшы да жойылады (65-сурет, 2). Бүл жағдайда оның ашық ұштары митозда шеңбер тәрізді хромосома түзе отырып қосылуы мүмкін.
Хромосоманың бір иіні бір мезгілде екі жерден үзілгенде де хромосома бөліктерінін, жетіспеушілігі байқалады (65-сурет, 3). Үзілген жерлер қосылады да, хромосома қысқарады, бүл кезде ішкі учаске жойылады. Егер бөлініп қалғаи үзінді айтарлықтай үлкен болса, жойылар алдында оның ашық үштары қосылуы мүмкін, ал метафазада ацентрлік шеңбер түзіледі.
Хромосоманын, шағын учаскелерінің бөлініп қалуына байланысты микроделецияны шамалы жетіспеушілікке жатқызады. Ұсақ жетіспеушіліктер кейде фенотиптік эффект бере отырып және гендік мутацияны бейнелей отырып әдетте гомозиготалы күйде сақталады. Оларды ген мутациясынан айыратын бірден-бір көрсеткіш—оларда кері мутациялану болмайды.
Хромосома бәліктерінің үлкен жетіспеушілігі әдетте гомози-готалы күйде летальды болады, бұл хромосоманың әрбір локусының клетка мен организм тіршілігі үшін зор маңызы бар екен-дігін дәлелдейді. Жетіспеушіліктен гетерозиготалар ғана тірші-лікке қабілетті болуы мүмкін. Бұл жағдайда жетіспеушілік арқылы анықталатын мутациялар доминанттық ретінде көрініс береді- Хромосома учаскесі жетіспеушілігінің фенотиптік эффек-тісі хромосоманың генетикалық системасын, гендердің орналасу реттілігін, олардың өзара байланысын бұзатындығымен түсін-діреді.
Ірі жетіспеушіліктерді генетикалық және цитологиялық әдіс-термен анықтауға болады. Мәселен, адамның «мысықша мия-улауының» синдромы сипатталған. Бұл синдром баланың сәби кезіндегі мысықтың мияулауына үқсас ерекше жылайтынына байланысты осылайша аталған. Ақыл-ой және дене кемістігі, микроцефалия және басқа бірқатар аномалиялар осындай сипат-ты белгілер болып табылады. Цитологиялық зерттеулер арқылы 5-хромосоманың біріндегі қысқа иіннің шамалы интеркалярлық делециясын табады. Бір шеңберлі 5-хромосомасы бар ауру адамның осындай аномалиясы сипатталған. Бұл жағдайда шең-бер түзуде де 5-хромосоманың қысқа иінінде делеция байқала-ды. Адам, сондай-ақ жануарлар мен өсімдіктер үшін де мүндай мысалдар кептеп сипатталған.
Егер қалыпты жағдайда әрбір хромосомада ген бір дозамен берілсе, онда ол екі еселенгенде немесе көбейгенде—дуиликация-да— геннің дозасы тиісінше 2 есе және одан да артады. Мысалы, егер қалыпты хромосомадағы гендер АВС тәртібімен орналасса, гендердің бірінің дупликациясы кезінде АВВС немесе АВВВС күйі пайда болуы мүмкін және т. с. с. Сірә, қүрамында бірнеше ген болатын хромосоманың үқсас учаскелерінің қайталанып ке-луі, мысалы, адамнан, тышқаннан, нейроспорадан, аспергилден, жүгеріден және т. б. табылған АВСАВСАВС дупликацияның не-ғүрлым жиі кездесетін жағдайы болса керек.
Дупликацияланған учаскелер хромосоманың көршілес учаскелерінде ғана болып қоймайды, сонымен қатар, бүкіл хромосомаға таралуы, ал кей жағдайларда басқа хромосомаларға да орын ауыстыруы мүмкін. Сірә, ұқсас учаскелердің көбеюі хромосомалар мен түрлер зволюциясындағы кең таралған қүбылыс болар.
Егер учаскелердің бірінде гендер АВСД тәртібімен қалыпты орналасса, олардың ВС учаскесін 180°-қа айналдыру инверсия-сында бүл тәртіп АСВД болып өзгеруі мүмкін.
Хромосома ішінде инверсия түзілуі үшін ол екі жерден үзілуі қажет. Инверсия көбіне рецессивті летальды эффектімен байла-нысты, сондықтан олар гомозиготалы күйде сақталмайды және оларды әдетте гетерозиготадан табады.
Алайда летальды эффектімен байланысы жоқ инверсиялар да кездеседі.
Инверсияны алып хромосомалардан немесе мейоздьщ пахитена сатысында цитологиялық әдіспен табуға болады. Гомози-готалы инверсияда хромосомалар коньюгациясы мен кроссинговер қалыпты жүзеге асады. Гетерозиготалы инверсияда хромосомалар коньюгациясы 66-суретте көрсетілгендей тәртіппен өтеді. Инверсияланған хромосома ілмек түзеді, ал қалыиты хромосома оны айналып өтеді.
Егер хромосомалар арасында алмасу бір рет өтетін болса, оның нәтижесінде әдеттен тыс: біреуі клетка бөлінуі кезінде жойылатын центромерсіз, ал екіншісі—екі центромері (дицентр-лі хромосома) бар екі хромосома түзілетінін 66, 3-суреттен көру-ге болады. Мұның кейінгісі I анафазада центромерлер полюстерге қарай ажырағанда хромосома «көпірін» түзеді. Бұл көпір кез келген жерден үзілуі мүмкін, осының нәтижесінде гаметалар бір учаскелерінде азды-көпті жетіспеушіліктер бар және екінші учаскелерінде дупликациялары бар хромосомаларға ие бола алады, сөйтіп тіршілікке қабілетсіз болып шығады. Тіршілікке қабілетті қалыпты гаметалар айқасуға түспеген немесе қос кроссинговерден өткен хромосомалар есебінен ғана түзілуі мүмкін (66-сурет, 4). Алайда соңғы жағдай өте сирек кездеседі. Өйткені кроссоверлі гаметалар дара кроссинговер есебінен тіршілікке
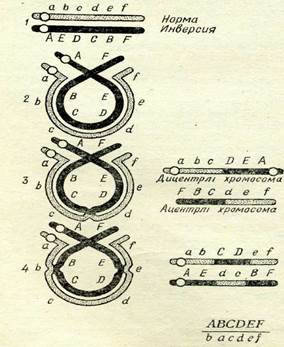 |
66-сурет. Акроцентрлік хромосомадағы гетерозиготалы инверсия жағдайындағы коныогация мен кроссинговер және мейоз өнімдері:
1 — ңальшты (аЪсйеі) және коныогацияға дейінгі ин-версияланғаи (АЕВСВҒ) хромосомалар; 2 —• коныогация; 3 — бір реттік крос-синговер жөне мейоз өнім-дері (дицентрлі және ацен-трлі хромосомалар); 4 — ңос кроссинговер және мейоз өнімдері: центро-мерлер ашың дөцгелекиен белгіленге.
кабілетсіз болып шығатындыктан, инверсия бойьшша гетерозиготаларда кроссинговердің болмайтындығы немесе оның тұн-шықтырылғаңдығы жөнінде әсер туады. Сондықтан инверскя-ларды кейде кроссинговерді бекітушілер деп атайды.
Инверсиялар жануарлар мен өсімдіктердің табиғи популя-цияларында кездеседі, олар адамда да ұшырасады. Адамда ге-терозиготада инверсияның болуы нормадан ешқандай ауыткушылықты туғызбауы мүмкін. Алайда (66-суретте) айтылып өт-кендей гаметогенезде жетіспеушіліктері мен дупликациялары бар теңестірілмеген гаметалар пайда болуы мүмкін, демек ұрпақта да сондай кемістіктер кездеседі.
Хромоссма аралық өзгерістер. Қарастырып өткен хромосомаішілік қайта құрылудан басқа, гомологты емес хромосомалар арасында учаскелер алмасумен байланысты езгерістер де болады. Хромосома аралық мұндай қайта құрылуды
Қалыпты хромосоманың бір жұбында жағдайда орын ауыстырған учаскелердің үзындықтары тең н-месе тең емес болуы мүмкін, сейтіп олардағы гендердің мөлшері де тең немесе тең емес болуы мүмкін. Алмасудың мүндай типін өзара немесе реципрокты транслокация деп атайды.
Транслокацияның басты генетикалық эффектісі ілінісу тобы-ның өзгеруі болып табылады: орын ауыстырған гендер жаңа тіркесу тобына енеді де, осы арқылы генотиптің қалыптасқан системасы бүзылады.
Мейоздағы транслокацияланған хромосомалар әрекетінің зор маңызы бар, өйткені гетерозиготалы особьтардағы мүндай хромосомалардың коньюгациясы өзінше ерекше етеді. Транслкация бойынша гетерозиготаларда I профазадағы коньюгация кезінде крест сияқты фигура пайда болады. Ондай фигуракын, пайда болу себебі әр хромосомаларға белініп кеткен гомологты локустар зигонема стадиясында бір-біріне өзара тартылысуға душар болады (67-сурет). Крест тәрізді фигуралар диплонема стадиясында күрделі хиазмалар түзеді. Диакинезде хиазмалар центромерлерден хромосомалардың шеттеріне қарай сырғуы нә-тижесінде сақина тәрізді фигуралар түзіледі. Кейде хромосома-лар сақинасы аударылу арқылы сегіздік тәрізді фигуралар тү-зеді. Хромосомалардын, дәл осылай орналасуы тіршілікке қабілетті, теңестірілген гаметалар береді (67-сурет, /), өйткені бүл жағдайда.өзгерген хромосомалардың екеуі де немесе өзгермеген екі хромосомалар полюстің біріне қарай кетеді. Хромосомалар мейозда сақина тәріздес орналасқанда теңестірілмеген геномдры бар гаметалар түзіледі: олардың біреулерінде—гендер екі рет
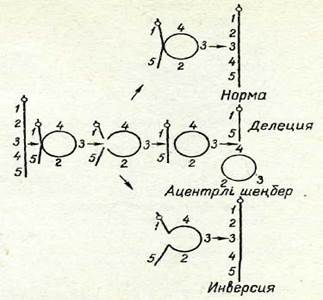 |
| 68-сурет. Акроцентрлі хромосомада үзілу пайда болған жағдайдағы хромосома ішілік қайта құрылудьң пайда болу механизмдері. Гендер цифрлармен белгіленген. |
Метацентрлі хромосомада да осындай әдіспен қалыпты құрылым қал-пына келуі немесе хромосомалық аберрация тууы мүмкін.
Хромосоманын. қызметін бірлеген жіпше (ерте интерфаза) немесе екі хроматид (кеш интерфаза және I профаза) атқаратын уақытта үзілу мен алмасу жүзеге асуы мүмкін. Бірлеген жіпше сатысында өтетін қайта құрылуды кейде хромосомалық қайта құрылу деп, ал екі хроматид сатысында жүзеге асатын қайта құрылуды хроматидтік қайта құрылу деп атайды.
Хромосомалық қайта қүрылуды зерттеу генетиктерге генотипті система ретінде зерттеу әдісін берді. Хромосомалар тұңым куалайтын дискретті, яғни олардьң әр түрлі локустары организмнің әр түрлі белгілері меи қасиеттерінің дамуын анықтайтын болса, әрбір хромосома езара әсер ететін гендердің эволюция процесінде қалыптасқан біртұтас система-сын көрсетеді. Инверсиялар мен транслокациялардағы гендер тәртібінің езгеруі көп ретте жаңа белгілер тууының себебі болатындығын көрсететін фактілер осы айтылғандарға дәлел болады. Қейбір дупликациялар мен жетіспеушіліктерде де осындай құбылыс байқалады.
Геннің басқа гендер системасындағы жағдайына байланысты оның әсерінің өзгеруін А. Стертевант жағдай эффектісі деп атауды ұсынды. Бұл генді дербес бірлік ретінде емес, генотиптің бүкіл системасынын, бір бөлігі ретінде қарастырудағы алғашқы талпыныс еді. Қазіргі кезде мүндай көзқарастық дүрыстығын дәлелдейтін көптеген фактілер бар (24-тарауды қарақыз).
Хромосомалық қайта қүрылудын, жәрдемімен генотиптердің жаңа системалары түзіле алады. Мәселен, тіршілікке қаблетті, транслокациясы, инверсиясы немесе дупликациясы бойынша гомозиготалы формалар пайда болатын жағдайларда ол белгілі бір тіршілік ету жағдайларына бейімделген болып,көбейіп, одан соң жеке түрге оқшаулануы мүмкін. Бүл жаңа түрдін, бүрынғы гендері сақталады, бірақ олар не басқа тіркесу тобында болуы керек немесе олардың хромосомада орналасу реті өзгеруі керек. Демек, хромосомалық қайта құрылу эволюция үшін де маңызды роль атқарады.
4. ГЕНОМДЫҚ МУТАЦИЯЛАР
Генамдық мутациялардың мэні және пайда болу себептері.
Әрбір түр үшін хромосоманың саны, формасы мен мөлшері сис-тематикалық белгілер болып табылады. Кариотиптің негізгі бір-лігі хромосомалардың гаплоидты жиынтығы, яғни гомологты хромосомалардың әрбір жүбынан біреуі ғана бар жиынтық. Мүндай гаплоидтық жиынтықта шоғырланған гендер тобын геном деп, ал гаплоидтық жиынтықтағы хромосомалар санын негізгі сан деп атайды да, оны п әрпімен белгілейді.
Митоз бен мейоз үрпақтан ұрпаққа берілетін хромосомалар санының түрақтылығын қамтамасыз ететін клетка бөлінуінің аса дәл механизмі болып табылады. Алайда кейбір жағдайларда бұл механизм бүзылады, ол хромосомалардың клетканың полюстері-не қарай әркелкі ажырауынан— хромосомалардың ажырамау-нан (8-тарауды қара), сондай-ақ хромосомалардың цитокинезсіз (эндомитоз) екі еселенуінен байқалады. Мүндай бүзылудың нә-тижесінде хромосомалар саны өзгерген клеткалар пайда бо-лады.
Түтас гаплоидты жиынтық санының немесе жекелеген хромо-сомалардық артуы немесе кемуі есебінен хромосомалар саны өзгеруі мүмкін. Гаплоидты жиынтықтары түтас көбейген организм-дерді полиплоидтар деп атайды. Хромосомаларының саны гап-лоидты жиынтыққа еселі болмайтын организмдерді анеуплоидтар немесе гетероплоидтар деп атайды.
Организм клеткаларындағы хромосомалар санының езгеруі оның белгілері мен қасиеттерінің өзгеруімен қатар жүреді, сон~ дықтан оларды геномдық мутациялар деп атайды.
Полиплоидия. Полиплоидия — гаплоидты жиынтыққа қара-ғанда еселенген хромосомалар санына артатын геномдық мута-циялар. Хромосомаларынын, гаплоидты жиынтықтарыньщ саны әр түрлі болып келетін клеткаларды: Ъп —триплоидты, Ап —те-траплоидты деп және т. с. с. атайды. Полиплоидты клеткалардан дамып жетілген организмдерді тиісінше триплоидтар, тетрапло-идтар деп атайды және т. с. с.
Гомозиготалы организмнен пайда болған тетраплоид та го-мозиготалы болады. Егер жиынтықтың кебеюі гомологты хромосомаларында белгілі бір гендердіқ әр түрлі аллельдері болатын буданды организмде жүзеге асатын болса, онда түзілетін: тетраплоид та осы гендер бойынша гетерозиготалы болады.
Соматикалық клеткалар полиплоидизацияланған жағдайда клеткалар организмнің бастапқы полиплоидтық клеткадан дамыған белігінде ғана полиплоидты болады да, организм химер-лы болып шығады. Егер полиплоидизация
зиготаның бірінші бе-лінуінде жүрсе, онда ұрықтың барлық клеткалары 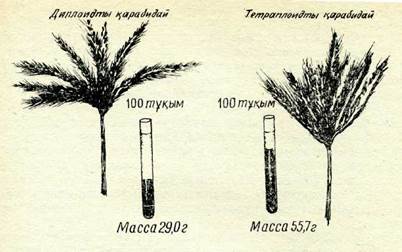
69-сурет. Сол жақта диплоидты және оң жақта тетраплоидты қара бидай.
болып шығады. Барлық хромосомалардың мейозда ажырамауы хромосомаларының саны редукцияланбаған гаметалардың түзі-луіне әкеп соғады, мұндай гаметаларда хромосомалар жиынты-ғы бірден емес, екіден болады. Хромосомаларының жиынтығы редукцияланбаған гаметалар ұрықтануға қатысқанда полипло-идты организмдер пайда болуы мүмкін.
Полиплоидия организм белгілерінің өзгеруіне әкеп соғады, сондықтаи да ол эволюция мен селекцияда, әсіресе өсімдіктер эволюциясы мен селекциясында өзгергіштіктің маңызды көзі бо-лып табылады. В. С. Федоров селекциясындағы диплоидты және тетраплоидты қара бидай өсімдігінін, фото-суретінде бұл жақсы керінеді (69-сурет). Тетраплоидты форманың сабағы, масағы «дәуір қуатты және дәні ірі болып келеді, бұл шаруашылық тұр-ғыдан алғанда оны барынша тиімді етеді.
Бидай туысы (Тгііісит) бірнеше турден тұрады, олар хро-мосомаларының саны бойынша, еондай-ақ өсімдіктің қасиеттері мен белгілері жөнінен үш топқа бәлінеді. Бірінші топқа, мыса-лы, соматикалық клеткаларында саны 14-ке тең диплоидты хро-мосомалары бар бір дәнектілер (Т. топососсит) жатады. Екін-ші топқа 28 хромосомасы бар қатты бидай (Т. сһігигп) жатады. 42 хромосомасы бар жүмсақ бидай (Т. аезііүит) үшінші топқа кіреді. Егер бидайда хромосоманың негізгі еа-ны п = 7 болса, бір дәнектілер диплоидты (7X2=14), қатты бидайлар—тетрапло-идты (7X4 = 28), ал жүмсақ бидайлар—гексаплоидты (7X6 = 42) болып шығады. Сүлы (Аұепа) туысы ішінде және басқа көптеген өсімдіктерде полиплоидтардың осындай қатары бел-гілі.
Хромосомаларынын, жиынтығы негізгі санға еселенген, хро-мосомаларының саны еселеніп артып отыратын қатар құрайтын туыстас турлгр тобын полиплоидтық қатар деп атайды. Поли-плоидтық қатар екі мүшелі және көп мүшелі болуы мүмкін.
Автополиплоидия. Бір түрдің геномдарының көбеюі негізінде пайда болған полиплоидтарды автополиплоидтар деп атайды. Егер хромосомалардың негізгі санын (геном) А әрпі-мен белгілесек, онда А гаплоидқа, АА —автодиплоидқа, ААА— автотриплоидқа, АААА — автотетраплоидқа сәйкес келетін бо-лады және т. с. с.
Автополиплоидтар табиғи жағдайларда кез келген әдіспен көбейетін организмдерде пайда болады. Автополиплоидтар әсі-ресе апомикті және вегетативті жолмен көбейетін формаларда өте бағалы, өйткені олар салыстырмалы алғанда өзгеріссіз түр-де үзақ уақыт бойы сақтала және көбейе алады. Егер бастапқы формасы гетерозиготалы болса, жыныстық жолмен кебеюде ав-тополиплоидтар түқым қуалаушылығы алуан түрлі формалар береді.
Автополиплоидтар мейозының диплоидтар мейозынан айыр-машылығы бар. Мәселен, тетраплоидта профазада биваленттер ғана емес, сонымен қатар (барлық гомологты хромосомалар өза-ра коньюгациялана алатындықтан) триваленттер, квадривалент-тер және униваленттер де түзіледі. Плоидтылық едәуір жоғары болғанда барлық гомологты хромосомалардың коньюгацияла-нуы поливаленттердің немесе мультиваленттердің түзілуіне әкеп соғады.
Егер диплоидты организм хромосомаларының бір жүбы бел-гілі бір ген (Аа) жәнінде гетерозиготалы болса, мейоз нәтиже-сінде гаметалардың екі сорты 1 А: 1 а пайда болады. Автотет-раплоидтын, ААаа гетерозиготалы диплоидтан пайда болған ре-дукциялық белінуінде гомологты хромосомалар полюстерге мы-надай қатынастарда ажырауы мүмкін: 2: 2, 3: 1, 1: 3, 4: 0, 0: 4. Осы жүптың үш, бір хромосомалы және хромосомасыз гамета-лары, атап айтқанда ААа және а, Ааа және А, сондай-ақ О то-лық бағалы болып есептелмейді. Бүл тіршілікке қабілетсіз зи-готалардың пайда болуына әкеп соғады, яғни полиплоидтардың фертильділігін кемітеді.
Бірақ тіпті гетерозиготалы ААаа автотетраплоидта хромосо-малар полюстерге қарай үнемі 2: 2 қатынасында тарайтын бол-са, тетраплоидтардағы ажырау бүл жағдайда да диплоидтардын. моногибридті ажырауынан өзгеше болады. Берілген аллель бо-йынша гетерозиготалы ААаа автотетраплоид гаметалардын \АА: ААа: \аа қатынасындағы үш типін түзеді: Ғ2. дефенотип бойынша ажырау 35: 1 болып шығады, яғни осы секілді дипло-идтан (3: 1) айтарлықтай өзгеше болады. 35: 1 қатынасындағы ажырау автотетраплоидты өсімдіктерге жүргізілген тәжірибелер арқылы әлденеше рет дәлелденді, атап айтқанда, ол түңғыш рет гүлдің күред қызыл және ақ бояуының түқым қуалаушылығын зерттеуде сасық меңдуанамен (Даіига) жасалған тәжірибелер-ден алынды. Диплоидтардың 1/2-і ғана гетерозиготалы болатын, болса, Ғ2-нің 36 буданыиың 34-і гетерозиготалы болуы айтарлықтай маңызы бар жағдай. Гексаплоидта /Үде ажырау 399: 1
болады.
Бұдан диплоидияға қарағанда полиплоидия гетерозиготалық-ты едәуір дәрежеде сақтайды деген қорытынды шығады.
Автополиплоидтар генетикасын зерттеудің ерекше маңызы бар, өйткені оларда рецессивті және доминантты аллельдердің арақатынасы бастапқы диплоидтардағыдай болып сакталады, ал белгілері өзгереді. Бұл плоидтылықтың белгілер көрінісіне әсерін зерттеуге мүмкіндік береді. Полиплоидтардың бәоіне ор-тақ қасиет клеткалар көлемінің өсуі болып табылады.
Клеткалардың көлемінің өсуі көбіне олардың бірқатар фи-зиологиялық және биохимиялық қасиеттерінің өзгеруіне байла-нысты: су мөлшерінің артуы, осмостық қысымның кемуі, әр түрлі заттардың өзгеруі белоктардың, хлорофилдің, клетчатканың, ауксиннің, бірқатар витаминдердің және т. б. Заттардың мөл-шерінін, өзгеруі қосымша құбылысты туғызуы, мысалы, поли-плоидтардың сыртқы факторлардың тербелісіне, ауруларға тө-зімділігін өзгертуі мүмкін және т. с. с. Полиплоидтардың едәуір жоғары адаптациялық қасиеттері, сөзсіз, олардың түқым қуа-лаушылығын байытуымен, яғни едәуір жоғары гетерозиготалы-лығымен де анықталады. Ежелгі өсімдіктер топтарының өкілде-рі — «тірі қазбалар» өздерінің кейінірек шыққан туыстарына қарағанда көп ретте полиплоидты болып шығатыны қызық жағ-дай.
Аллополиплоидия. Әр түрлі түрлер геномдарының көбеюі нәтижесінде пайда болатын полиплоидтарды аллополи-плоидтар немесе амфиплоидтар деп атайды. Мысалы, түр ара-лық буданда А және В геномдары қабысатын болса, одан алын-ған аллотетраплоид ААВВ болады. Аллополиплоидияны басқа-ша түрде будандық полиплоидия деп атайды. Оның зор практи-калық маңызы бар.
Алшақ будандар көп ретте үрықсыз болып шығады (мыса-лы, қара бидайдың бидаймен, шомырдың капустамен буданы және т. б.). Бүл қүбылыстың себептерінің бірін қарастырайық. Шомыр мен капустаның геномдары)? және В қабысты делік, бүл жағдайда шомыр-капуста буданында әр түрлі екі геном— Я және В болады. Шомыр будан зиготасына 9 хромосомадан тү* ратын геномын, капуста да 9 хромосомадан түратын В геномын енгізеді. Мүндай алшақ Ғ\ буданының соматикалық клеткала-рындағы хромосомалардың жалпы саны 18. Мейоз профазасын-да жыныс клеткаларының түзілуі процесінде гомологты хромо-сомалар коньюгациясы жүруі тиіс.
Алайда шомыр хромосомаларының капуста хромосомалары ішінде гомологтары болмайтындықтан, мейозда олардың әрқай-сысы өзін унивалент ретінде жеке үстайды. Мейозда көрсетілген будан клеткаларынан 18 унивалентті санауға болады. I анафа-зада олар полюстерге қарай тәртіпсіз таралады да, осының нә-тижесінде хромосомаларының саны 0-ден 18-ге дейінгі түрліше шамада кездесетін гаметалар түзіледі. Олардың кәпшілігі теңестірілмеген болып шығады, яғни олардағы хромосомалардың саны нёгізгіге қарағанда еселенбеген, сондықтан тіршілікке қабілетсіз болып шығады.
Алайда мұндай будандағы аналық, сондай-ақ аталық гаметалардың кейбір бөлігінде 9К + 9В = 18 хромосома болады. Бүл гаметаларды редукцияланбаған деп атайды. Редукцияланбаған гаметалардың ұрықтану процесінде бірігуінен екі түрдің де хро-мосомалар жиынтығы екі еселенген зигота—аллотетраплоид немесе амфидиплоид пайда болады. Онда шомыр хромосомалары-ның екі жиынтығы (9К + 9В) және капуста хромосомаларының екі жиынтығы (9К + 9В), яғни барлығы 36 хромосома болады және оның мейозында әрбір хромосоманың коньюгацияланатын серігі болатындықтан фертильді болып шығады.
Амфидиплоидтарды алу будандастыру жолымен және будандардың хромосомалар санын екі еселендіру жолымен жаңа константтық формалар синтездеу мүмкіндігін ашты, өйткені олар бастапқы формаларды бермейді және өздеріндегі констант бел-гілерге аралық түқым қуалаушылық тән болады. 20-жылдардың басында Г. Д. Қарпеченко шомырды (Карһагшз заііуиз) капус-таман (Вгаззіеа оіегасеа) будандастыру арқылы алғаш рет туыс аралық өсімтал будан алды. Бүл өте мықты будан болып
шықты, әрі ол шомыр мен капустаның белгілерін біріктірді. Ол келесі ұрпақ-тарда да өсімтал әрі константты күйінде қалды. Екі туыс геномдарының үйлесуі негізінде синтезделген бүл жаңа форма рафанобрассика (КарһапоЬгаззіса) не-месе шомыр-капуста буданы деп аталды. 70-суретте рафанобрассиканын (4) жемісі мен хромосомалар жиынтығы кел-тірілген. Бұл форманың бүршаққынының: жоғары бөлігі шомырдан (70-сурет, 1), ал оның табаны капустадан (70-сурет, 2) қүрастырылған болып шықты. Бүл
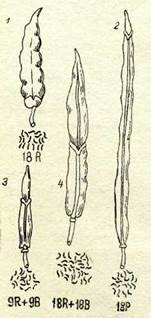 |
| 70-сурет. Шомырдьщ, капустаның және олардьң будандарының жемістері мен хромосома жиынтықтары: |
буданың диплоидты формасы ұрықсыз бо-лады (70-сурет, 3).
Әр түрлі геномдарды үйлестіру және полиплоидизация жолымен табиғатта жоқ жаңа формаларды синтездеуге болатыны қарастырылған мысалдан көрінеді. Мұндай жаңа формаларды шығаруды түрлердің синтезі деп атайды. Бірқатар ұрпактар бойы сүрыптаудан кейін олар толығымен константты бола алады және оларды дербес таксономиялық бірліктер деп санауға болады.
.Полиплоидтар туралы айтылғандардан, олардын, өсімталдығы гомологты хромосомалардың мейозда коныогациялану сипатымен анықталады деген қорытынды шығады. Алайда хромосомалар цитоплазмамен тығыз әрі күрделі байланыста болады. Клеткалар бірыңғай система ретінде қызмет атқарады, сондықтан полиплоидтар өсімталдығындағы айырмашылық кейде геномдардың цитоплазмамен сыйысатындығымен немесе сыйыспайтындығымен анықталуы мүмкін.
Цитоплазманың аллополиплоидтар гаметогенезіндегі ролін рафанобрассика мысалъшда көрсетуге болады. Будан шомыр гүлін капуста тозаңымен тозаңдандырудан алынғандықтан, бұл форманын, цитоплазмасы шомырға жатады. Цитоплазманын ро-лін анықтау үшін Г. Д. Карпеченко екі ата-аналық формалар-мен кері будандастыру жүргізді. Ғ будандарында «шомыр» цитоплазмасы ғана болды, ал хромосомалардың арақатынасы өзгеріп отырды. Бірінші шағылыстыруда рафанобрассикаХшо-мыр аллотриплоидының мейозда шомыр хромосомаларының 9 биваленті және капуста хромосомаларының 9 униваленті бол-ды (9і?4-9і? + 9В). РафанобрассикаХкапуста екінші будандас-тырылуында хромосомалардың кері қатынасы болды, бірақ екі жағдайда да 9 бивалент пен 9 унивалент түзіл-ді. Демек, екі триплоидтың да мейоз барысымен өсімталдығы бірдей болуы тиіс деп күтуге болатын еді. Шындығында да,. сырт қарағанда мейоз буданның екеуінде де бірдей өтетін болып шықты, бірақ екінші жағдайда гаметалар толығымен қырыла-ды, ал бірінші жағдайда—қырылмайды- Мұнын, себебі мынада:: бірінші жағдайда шомырдың 9 биваленті үшін шомыр цито-плазмасы өзінікі, ал екінші жағдайда—капустаның 9 биваленті үшін бөтен болып шығады.
Сонымен, цитоплазмасы бірдей, бірақ хромосомаларынын жиынтығы түрліше аллополиплоидтардың өсімталдығы жөнінен айырмашылығы болады., өйткені аллополиплоидтар гаметогене-зінде цитоплазма айтарлықтай роль атқарады.
Полиплоидия өсімдіктерде кеңінен тараған. Бұл оларда гермафродитизмнің (өздігінен тозаңдануы), апомиксистің (парте- ногенез) және вегетативтік көбеюдің барынша кеңінен таралғандығына байланысты. Сірә, өсімдіктерде полиплоидияның жүзеге асуына себепші болатын басқа да себептер болса керек.
Жануарлар полиплоидиясы. Дара жынысты жануарларда полиплоидия өте сирек кездесетіні анықталды.
Қазіргі кезде түт көбелегінің, тритонның, аксолотльдің автополиплоидты особьтары алынды. Жібек құрты (ВотЬух тогі) автотетраплоидтарының ұрғашылары есімтал, ал еркектері стерильді келеді. Мүның себебі, еркектерінде (гомогаметалы жы-ныс) мейоз профазасында поливаленттер түзіледі, осының нәти' жесінде хромосомаларының саны анеуплоидты болып келетін тіршілікке қабілетсіз гаметалар пайда болады. Ұрғашыларының (гетерогаметалы жыныс) хромосомалар коньюгациясы шектел-ген, поливаленттер түзілмейді, ал пайда болушы гаметалар тір-шілікке қабілетті келеді. Дара жынысты жануарларда полиплоидтардын. сирек кездесу себептерінің бірі — олардың ұрық-сыздығы.
Жыныстық көбе.юі партеногенезбен алмастырылған жануарларда полиплоидия түгелдей дерлік өсімдіктердегідей жүзеге асуы мүмкін.
Жануарлардағы полиплоидтық қатар мысалына олардың аскаридадағы, жер қүртындағы, қос мекенділердегі, көбелектер-дегі және басқалардағы көрінісін жатқызуға болады.
Сүт қоректілерден триплоидты зиготалар табылған. Алайда триплоидтық эмбриондар тышқандарда эмбриондық дамудың ортасына дейін ғана тіршілік етеді. Адамда спонданды жасалатын аборттар немесе баланын, өлі тууы материалдарын талдау нәтижесінде триплоидия және тетраплоидия жағдайлары табылған.
Жануарлар аллополиплоидтарын жасанды жолмен қолдан алу соңғы уақытқа дейін шешілмейтін проблема болып есептеліп келеді. Алайда таяуда қызметкерлерімен бірге Б. Л. Астау-ров ВотЪух тогіХВ. тапсіагіпа жібек көбедегінің түр аралық буданынан алғашкы аллополиплоид алды. Аллотетраплоидты синтездеуде жасанды партеногенез әдісі пайдаланылды. Шағылыстыру процесінде екі жыныстьщ да әр түрінің хромосомалар жиынтығы екі еселенген формалары сұрыпталып алынды: 1п В. тогі + 2/г В. тапсіагіпа — аллотетраплоидтар немесе амфидиплоидтар.
Жануарлар дүниесіндеғі полиплоидия бірқатар тканьдердің соматикалық клеткаларында, яғни митоз жолымен мейозсыз көбейетін жағдайларда айтарлықтай кең таралған.
Қазіргі кезде генетиктердің көпшілігі жануарлар эволюция-сында полиплоидия емес, хромосома аралық және хромосома ішілік қайта қүрылулар негізгі роль атқарады деген көзқарасты жақтайды.
Анеуплоидия. Анеуплоидия (немесе гетероплоиаия) дегеніміз гаплоидты саны еселенбеген, хромосомалар санының өзгеруінен түратын геномдық мутация.
Бүл қүбылысты дрозофиланың жыныспен тіркескен белгілерінін, түұқым қуалауын зерттеуде К- Бриджес алғаш рет таза ге-нетикалы.қ әдіспен анықтады. Дрозофиланың ұрғашысында жы-ныс хромосомаларының тарамауы жағдайында белгілердің тұқым қуалау мысалдары 8-тарауда келтірілген.
Жекелеген хромосомаларының саны өзгерген клеткалардың пайда болуын клетка бөлінуде хромосомалардың кейбір жұптарыньң тарауы бүзылатындығымен түсіндіруге болады. Мүндай бұзылушылықтар соматикалық және жыныс клеткаларында кез-десуі мүмкін. Жоғарыда сез болған, кейбір полиплоидтар мен алшақ будандар секілді гомологты хромосомаларынын коньюгациясы бүзылған формаларда тұрақты анеуплоидты гаметалар пайда болады.
Егер қосымша хромосомасы (я+1) бар гамета қалыпты, гаплоидты (п) гаметамен үйлессе, зиготада бір хромосома артық болып шығады: ондағы хромосомалар саны 2я+1-ге тең бола-ды. БІр хромосомасынан айрылған гамета қалыпты, яғни гапло-идты гаметамен үйлессе, диплоид саны толық емес, бір хромосома жетіспейтін (2 п —1) зигота пайда болады.. Хромосомаларының жиынтығы 2«+1 болатын организмді трисомик деп, 2п —1 болатын организмді— моносомик деп, ал 2п —2 болатын организмді— нулисомик деп атайды. Кейбір си-рек жағдайларда хромосоманың белгілі бір жұбының қосымша бір хромосомасы емес, екі хромосомасы (2/г + 2) — тетрасомигі, үш хромосомасы (2/г+З)— пентасомигі болуы мүмкін және т. с. с.
Мүндай санның кемуі немесе артуы хромосомалардың кез келген жүбынан байқалуы мүмкін, сондықтан гомологты емес хромосомалардың бірнеше жүбында бір мезгілдік анеуплоидия болуы мүмкін.
Анеуплоидия белгілердің тұқым қуалау сипатының өзгеруіне ғана әкеп соқпайды, сондай-ақ ол фенотипте де белгілі бір өзгеріс туғызады. Мәселен, адамда жиынтықтың барлық хромосомалары бойыиша трисомия сипатталған. Трисомия кейде толық болады, яғни бір нөмірдің үш хромосомасы қайталанады, ал кейде—ішінара болады, мұнда екі хромосома толық, ал үшінші хромосома ішінара қайталанады. Трисомияның мүндай жағ-дайы әсіресе геномның ірі хромосомаларында жиі кездеседі. Бүл жекелеген хромосомалардың генетикалық қүндылығы әркелкілігін білдіреді. Ішінара трисомия негізінен алғанда инверсияның немесе геномдағы дупликацияның болуы есебінен түзіледі. Әр-бір хромосоманың трисомиясы фенотиптік түрғыдан алғанда симптомдардың белгілі бір жиынтығы болуымен сипатталады, бірақ бұл әрқашан да көптеген кемістіктердің жиынтығын қам-титын психомоторлы дамудың бүзылуы болады.
Бидай, жүгері, темекі, мақта, тышқан, мысық, ірі кара мал және т. б. анеуплоидтары сипатталған. Олардың тіршілікке қабілеттілігі әдетте нашар, тіршілік үзақтығы қысқа, диплоидтарға қарағанда өсімталдығы шамалы және диплоидтардан морфо-логиялық белгілері жөнінен көптеген айырмашылықтары болады. Жануарларға қарағанда өсімдіктер анеуплоидиясы олардың тіршілік қабілеттілігіне айтарлықтай әсер етпейтіні белгілі.
Анеуплоидтарда қалыпты гаплоидты, сонымен қатар анеуплоидты гаметалар түзіледі. Бүл жағдайда өсімдіктердің ұрықтануына хромосомаларының жиынтығы қалыпты, гаплоидты тозаңдар ғана қатысады, ал үрық қалталары хромосомалар са-нына тәуелсіз қызмет атқарады, сондықтан анеуплоидтар рпағындағы ажырау сипатынан өзгеше болады. Мысалы, беде өсімдігі хромосомасы женінен гүлдің қызыл(71) немесе ақ (а) гені бар трисомик болса, ААа генотипі кезінде өздігінен тозаң-данатын жағдайда ыдырау 17:1 болады. Мүны қызмет атқаратын екі сортты тозаң — А және а түзілетіндігімен түсіндіруге болады, бірақ мұнда а гені бар тозаң дәндеріне қарағанда А гені бар тозаң дәндері 2 есе көп болады. Мынадай пропорциямен:1 АА: \а:2А: ІАа төрт сортты (А, а, АА, Аа) жұмыртқа клеткалары пайда болады. Пеннет торы бойынша 17:1 қатынасын оңай алуға болады.
Қазіргі кезде генотиптегі әрбір хромосоманың ролін анықтауға байланысты өсімдіктер анеуплоидиясын зерттеудің зор маңызы бар. Болашақта бұл белгілі бір генотиптерді экспери-менттік жолмен синтездеуге жәрдемдеседі. Генотип эволюция-сында анеуплоидия зор роль атқарады және оның мәдени өсім-діктердің шығуын зерттеуде зор маңызы бар. Алайда бұл мәселелер алдағы уақытта мұқият қарастыруды талап етеді.
5. ЦИТОПЛАЗМАЛЫҚ МУТАЦИЯЛАР
Цитоплазмалық мутациялар үғымы.СОрганизмнің белгілері мен касиеттерінің өзгеруіне әкеп соғатын плазмогендердің өзгерісін цитоплазмалық немесе плазмалық мутациялар деп атайды. Цитоплазмалық мутацияларды зерттеу олардың орналасу орнын анықтаудағы нақты қиындықтарға байланысты күрделене түсті. Жоғарыда айтылғандай, қазіргі кезде плазмогендер пластидтер мен митохондрияларда ғана шоғырланған, ал басқа органоидтарда олардың болатындығы дәлелденген жоқ. \
Шоғырланған орны мүлде белгісіз плазмогендер де бар. Мысалы, жүгерідегі ІІМС-ін тек жобалап митохондрияда болады деп есептейді (10-тарауды караңыз).
Цитоплазмалық мутациялардың гендік мутациялармен ұқсастықтары бар: олар түрақты және үрпақтан үрпаққа беріледц Алайда оларды табу қиын, өйткені клеткада аттас цитоплазмалық құрылымдар көп мөлшерде кездеседі. Фенотипте рецессивті мутациялар байқалуы үшін клеткадағы тиісті цитоплазмалық қүрылымдардың бәрі мутантты болуы қажет. Мутантты және мутантты емес қүрылымдардың митозда бөліну жылдамдығы клеткадағы осы қүрылымдардьщ жалпы санына байланысты.
Цитоплазмалық мутациялардың табиғаты. Цитоплазмалық мутациялардың табиғаты алуан түрлі болуы мүмкін. Өзгерудің. екі типі белгілі. іЭнын, бірінші типіне қүрылымнан айрылу жатады. Қараңғы жағдайда үзақ уақыт өсіруден пластидтерінен айрылатын (пластидтік мутация) эвгленаның мутантты штаммалары осыған мысал бола алады. Құрылымы-нан айрылу қайтымсыз құбылыс болып табылады да, талшық-тылардың бірқатар касиеттерінің өзгеруіне себепші болады. Цитоплазмалық қүрылымдары атқаратын қызметі жағынан немесе морфологиялық түрғыдан өзгерген мутанттар екінші типке жатады. Бұған ашытқы мен нейроспорадағы «тыныс алу» мутациялары мысал бола алады (10-тарауды қараңыз). Мутантты және қалыпты клеткалардың электрондық микрофотографиялық суретінен олардың митохондрияларыныд морфологиясы жөнінен бір-бірінен айырмашылығы жоқ екені көрінеді. Ашытқының мутанттық штаммаларының митохондрияларында цитохромоксидаза болмайтындығы арнаулы зерттеулер арқылы керсетілген.Плазмогендер өзгерісі, сірә, митохондриялар ДНК-нің өзгерісімен байланысты болуы да мүмкін.
Мутантты плазмогендер рецессивті немесе доминантты бола алады. Олардың көрінісін доминантты плазмогеңдер басып тастайтын болса да, клеткада рецессивті плазмогендер сақталады және өздерін-өздері өндіреді.
Пластидтердің фенотипін анықтайтын цитоплазмалық мутациялар барлық жасыл өсімдіктерде спонтанды түрде пайда болады. Олардың туу жиілігі аса көп емес. Мысалы, энотерада шамамен 0,02%, есімдіктердің құрамында мутантты сары неме-се ақ пластидтері бар, яғни қалыпты жасыл бояуынан айрылған жекелеген клеткалары немесе ткань учаскелері болады. Шын-дығында да, мұндай мутациялардың пайда болу жиілігі едәуір жоғары, атап айтқанда 0,1%-тен 0,5%-ке дейін жететін өсімдік-тер (қызылша, капуста және т. б.) де бар. Кері цитоплазмалық мутациялардың пайда болу жиілігі мұқият зерттелмеген. Бірақ, кері мутациялану (аномальды пластидтерден қалыпты пластид-тердің тууы) фактісінің өзін анықталған деп есептеуге болады. Плазмогендердің ерекшеліктері. Кейбір химиялық заттар өңделетін клеткалардың көптеген мөлшерінде цитоплазмалық му-тациялар туғыза алады. Бұл тұрғыдан алғанда плазмогендердің хромосомалық гендерден түбегейлі айырмашылықтары бар. Хро-мосомалық гендерде осы агенттер әсер еткенде мутациялар 10~6 және одан да сирек жиілікте пайда болады.
Акридинді бояғыш заттар плазмогендерге әсер ететін химия-лық заттардың қатарына жатады. Мысалы, эуфлавин өңделген ашытқы клеткаларының кептеген мөлшерінде ергежейлі мутация туғызады.
Кейбір өте сирек жағдайларда сыртқы орта өзгерістеріне жауап ретінде пайда болатын мутациялар белгілі бір дәрежеде, атап айтқанда оларды туғызған жағдайларға адаптацияланғыш болып табылады. Мысалы, басқа мутациялармен қатар стрепто-мицин хламидомонаданың (Сһіатусіотопаз геіпһагсіі) сезгіш клеткалары ішінде төзімді, стрептомицин бар ортада тіршілік етуге қабілетті клеткалардың пайда болуына жағдай жасайды. - Сонымен, цитоплазмалық мутациялардың болуы түр генефондыи бүрынғыдан да байытып, эволюциядағы езгергіштіктің қосымша резерві болып қызмет етеді.
8. МУТАЦИЯЛАРДЫ ЕСЕПКЕ АЛУДЫҢ КЕЙБІР МЕТОДТАРЫ
Мутацияларды есепке алу методтарының ерекшеліктері.
Объектінің негізінен алғанда, организмнің кебею ерекшеліктері-не байланысты мутацияларды табу әдістері түрліше болуы тиіс. Мутацияның көріну сипаты да оны табу методын айқындайды. Кейбір көрінетін морфологиялық езгерістерді айтарлықтай дәл анықтауға болады: көп клеткалы организмдердегі физиология-лық және биохимиялық өзгерістерді анықтау едәуір күрделі бо-лып табылады. Көрінетін доминантты мутациялар бәрінен де оңай табылады, өйткені олар бірінші ұрпақтың өзінде-ақ гет-розиготалы күйде байқалуы мүмкін, ал рецессивті мутацияларды талдау қиындау болады, оларды гомозиготалы күйге келтіру қажет. Рецессивті мутацияны анықтау үшін бірқатар үрпаққа ар-найы генетикалық талдау жүргізу талап етіледі.
Генетикалық түрғыдан алғанда жақсы зерттелген, анықтал-ған тіркесу топтары бар объектілер (дрозофила, жүгері, бірқатар микроорганизмдер) үшін жаңа мутацияларды зерттеу жұмысын жүргізу айтарлықтай оңай болады. Бұл объектілер үшін мутациялар жиілігін есепке алудың арнаулы методы жасалған.
Дрозофиланың көрінетін мутацияларын есепке алу. Дрозо-филаның жыныс хромосомасындағы көрінетін мутацияларды табу үшін хіабайы типтің талданатын особьтарын Х-хромосомадағы рецессивтік гендер қатары бойынша гомонемесе гемизиготалы болып табылатын особьтармен шағылыстыруды пайдалануға болады. Бұл жағдайда Ғгден жабайы типтің гетерозиготалы
ұрғашыларымен І^г-Аг іқатар, белгілі бір рецессивті белгілері
көрінетін жекелеген особьтарды (==гзг) Да байқауға болады.
Олардың көрінуі тиісті генде (бұл мысалда а гені) мутацияньщ пайда болғанын білдіреді.
Дрозофиланың рецессивті, жыныспен тіркескен летальды му-тациясын есепке алу. Мутацияның мүндай типін есепке алу үшін Г. Меллер СІВ және М-5 (Меллер-5) методын жасады. М-5 ме-тодының нақты сипаттамасы «Генетиканың практикалық сабақ-тарына басшылықта» берілген. Бүл методтың мәні мынада: X- хромосомасында, гемизиготалы күйде рецессивті летальды мутациясы бар еркек особь тіршілікке қабілетсіз болады. Бұл жағдай.Гг-де фенотипі бойынша еркек особьтардың белгілі бір класының түзілмеуіне әкеп соғады, ал мүның өзі летальдардьщ бар екендігін көрсетеді-
Басқа объектілердегі мутацияларды есепке алу. Басқа объектілер (тышқан және т. с. с.) үшін де мутацияны табудың осындай методтары жасалған. Олардың негізіне де аталған принцип-тер алынған: 1) рецессивті мутацияны гомонемесе гемизиготалы күйге көшіру арқылы табуға болады, 2) гетерозиготалы особь-тарда кроссинговер болмайтын жағдайларда ғана мутациялардың пайда болу жиілігін дәл есептеуге болады.
Микроорганизмдер мутациясын табу методтары 15-тарауда қарастырылатын болады.
Доминантты летальды мутациялардың пайда болу жиілігін есепке алу методикасы сүт қоректілер (тышқан, үй қояны, ит, шошқа т. с. с.) үшін барынша қанағаттанарлық дәрежеде жасалды. Мутациялардың жиілігі туралы аналық безіндегі сары денелер мен іші жарылған буаз ұрғашының дамып келе жатқан эмбриондарының саны арасындағы айырмашылық бойынша қорытынды жасайды.
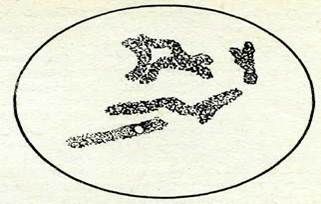 |
| 71-сурет. Сгеріз сарпіагіз тамыршасының клеткасындағы метафазадағы симметриялы хроматидті транслокация. |
Адамда мутацияның пайда болу жиілігін есепке алу өте қиын, алаида генеалогиялық, яғ-ни шығу тегі шешіресін талдау жаңа мутациялардын, пайда болуын анықтауға мүмкіндік бе-реді. Егер жұбайлардың шығу тегі шежіресінде бірнеше ұрпақ бойында қандай да бір белгі кездеспей, балаларының бірінен байқалса және келесі ұрпаққа тұрақты беріле бастаса, онда бүл жұбайлардың бі-рінің гамет.асында мутацияның пайда болғандығы жөнінде айтуға болады (бұл туралы 28-тарауда толығырақ айтылады).
Хромосомалық қайта құрылуларды есепке алу. Генетикалық методтармен табылатын летальды мутациялар генотиптік табиғаты бойынша әр түр- М болуы мумкін. Бүған әр түрлі хромосомалық қайта құрылулар, сондай-ақ жекелеген гендердегі өзгерістер жатады.
Белгілі бір типтегі хромосомалық қайта қүрылу жиілігін цитологиялық әдістерді пайдалану арқылы есепке алуға бола-ды. Дрозофила мен басқа да қос қанатты насекомдардың сілекей бездерінің алып хромосомалары талданады, ал өсімдіктердің немесе жануарлар мен адам тканінің культурасынан хромосомаларды митоз метафазасы мен анафазасында талдауға болады.
Анафазалық есепке алу методы көпірлер мен фрагменттерді анықтауға мүмкіндік береді. хЧетафазалық есепке алу методы еңбекті көп қажет етеді, бірақ дәл болады, ол зақымданудың басқа да түрлерін, мысалы үзілу мен алмасуды үқсастырып теңестіруге мүмкіндік береді. Симметриялы хроматидті транслокация кезінде сіңлілі хроматидтер арасындағы коньюгация әсері-нен метафазада крест сияқты фигуралар" пайда болады, ол 71-суретте жақсы көрсетілген.
Хромосомаларды дифференциалды бояу методы жануарлар мен адам тканьдерінің культурасындағы клеткалардан хромосо-малық аберрация типін айқындап қана қоймай, сондай-ақ хромосоманың қандай локустары қайта қүрылу процесіне ушырағанын анықтауға да мүмкіндік береді.
7. СПОНТАНДЫ МУТАЦИЯЛЫҚ ПРОЦЕСС ЖӘНЕ ОНЫҢ СЕБЕПТЕРІ
Мутациялар жиілігі және олардың себептері. Әдетте «спон-танды мутациялық өзгергіштік» терминіне әдеттен тыс агенттер-дің арнайы әеерінсіз табиғи жағдайларда пайда болатын мутациялар біріктіріледі. Мутациялық процесс негізінен мутациялар-дың пайда болу жилігімен сипатталады. Жануарлардың, өсімдіктер мен микроорганизмдердің әрбір түріне мутациялардың белгілі бір пайда болу жиілігі тән: бір түрлердің мутациялық өзгергіштіктігі басқаларына қарағанда едәуір жоғары болады.
Қазіргі кезде мутацияның бір үрпақта пайда болуының жалпы жиілігі женінде жеткілікті мәлімет жоқ. Мұны мутациялардың фенотиптік көрінісі жөнінен, сондай-ақ генетикалық табиғаты жағынан да барынша алуан түрлі болатындығымен, ал оларды есептеу методтары жетілмегендігімен түсіндіруге болады; жеке-леген локустардың мутацияланғыштығы жөнінде ғана белгілІ дәрежеде дәл баға беруге болады. Әдетте, аллельдік жұп мүшелерінің біреуі ғана бір мезгілде мутациялануы күмәнды жағдай.
Спонтанды мутациялану жиілігінің анықталған заңдылықтары мынадай қағидаларға келіп тіреледі: әр түрлі гендер бір генотипте түрліше жиілікпен мутацияланады (мутацияланғыш және тұрақты гендер болады); үқсас гендер әр түрлі генотиптерде түрліше жиілікпен мутацияланады.
Салыстырмалы түрде алғанда әрбір ген сирек мутациялана-ды, алайда генотиптегі гендер саны өте кәп болғандықтан, бар-лық гендердің мутациялануының жалпы жиілігі айтарлықтай жоғары болып шығады. Дрозофила ушін бір ұрпақта шамамен 100 гаметаға бір мутация келетінін, басқа организмдер үшін бұан да жоғары жиілік болатынын, мәселен, 10—30 гаметаға 1 мутация келетінін есептеу керсетті.
Селекцияның көмегімен әр түрлі спонтанды мутацияланғыш-тығы бар линиялар жасауға болады. Басқа гендердің мутацияла-ну жылдамдығына эсер ететін ерекше гендер —мутаторлардың болу фактілері де осы айтылғандарға дәлел болады. Мысалы, жүгеріде а генінің мутациялағыштығына әсер ететінгені бар. генотипі бар есімдіктерде антоцианмен боялмаған тканьдер фонында боялған учаскелер (а-дан Л-ға қарай сомати-калық мутациялар) пайда болады, ал сол уақытта өсім-діктерінде мүндай қүбылыс кездеспейді.
Спонтанды мутациялық процесс сондай-ақ клетканың физио-логиялық күйімен және биохимиялық әзгерістерімен де қамтамасыз етіледі. Мысалы түқымды бірнеше жыл сақтаудағы қар-таю процесінде мутациялардың, әсіресе хромосомалық қайта құрылу типіндегі мутациялардың жиілігі едәуір артатынын М. С. Навашин мен Г. Штуббе көрсетті.
Генотипте белгілі бір заттар биосинтезін тежейтін мутация-лардың жиналуы сп








