 2015-06-28
2015-06-28 6430
6430Бактериялардың белгілерінің түқым қуалаушылық сипатын зерттеуде де айтарлықтай қызықты мәліметтер алынды. Езсһегісһіа соіі бактериясында генетикалық информацияны бір бак-териядан екіншісіне тасымалдау клеткалардың коньюгация деп аталатын жанасуы процесінде жүзеге асатыны генетикалық және цитологиялық (электрондық микроскопия) әдістермен көрсетілді (VI таблица, А). Бүл берілісте тек бір жақты жүзеге асады. Бактериялық клетканың жалғыз хромосомасы түйық сақина пішінді болады, ол коньюгация кезінде ғана үзіледі және бір клеткадан екіншісіне өтеді.
Мұнда хромосоманың ролін ұзындығы 1,2—1,4 нм қос жіп-шелі ДНК атқаратынын ескертеміз. Берілген хромосомалық материалдың саны коньюгация үзақтығына байланысты, алай-да ол, әдетте бастапқы хромосоманың шамалы ғана бөлігі бо-лады. Сонымен, коньюгацияның нәтижесінде бір клетка түзіле-ді, оның түтас хромосомасы плюс басқа фрагменті болады. Мұндай клетка мерозигота деп аталады, онда генетикалық түр-ғыдан оңай талданатын рекомбинация процесі жүреді. Хромосо-маны беретін басқа клетка өзгеріссіз сақталады, өйткені коныо-гация кезінде ДНК репродукциясы жүреді.
Ішек таяқшасының бірқатар штаммаларын зерттеу олардың кейбіреулерінің арасында коньюгация жүрмейтінін, ал басқала-рының арасында ол рекомбинантты формалардың пайда болуы-мен ойдағыдай ететінін көрсетті. Бүл барлық штдммаларды екі топқа бөлуге және оларды жыныстық түрлер ретінде карасты-руға мүмкіндік берді, Олар Ғ+ немесе Ғ~ таңбасымен белгіле-неді. Әр түрлі жыныстық түрге жататын бактериялардың ша-ғылыстыру кезіндегі атқаратын қызметі де түрліше болатынын салыстыру керсетті. Ғ~ клеткаларының ішінен коныогациядан кейін 10~6 немесе 10~9 клеткаға 1 жиілікте мерозиготалар мен рекомбинанттар пайда болады. Сондықтан Ғ~ клеткаларын «аналық» клеткалар деп атайды. Ғ+ клеткасы ұрықтандырушы («аталық») клетка ролін атқарады, ол коньюгация процесіиде тұқым қуалау информациясын сақтайтын ДНК-ны береді.
Коньюгациядан кейін Ғ~ көптеген клеткалары культураныя басқа ешқандай белгілерін қабылдамастан, Ғ+ клеткаларынын. сипатына ие болатыны байқалған. Жыныстық фактордың бері-луі басқа генетикалық маркерлерге тәуелсіз етеді. 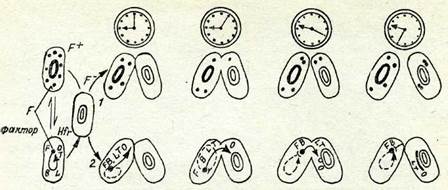
78-сурет Езсһегісһіа соіі-де (2) коныогацияның ұзақтығына байланысты генетикалық материалдьщ берілу тәуелділігі және Ғ-фактор (1) мысалында эпи-сомалардын. берілуі:Ғ+-«аталың» клеткалар; Ғ~ — «аналық» клеткалар; Щг —рекомбинацияныңжоғары аныңтайтын жыныс түрі: В, £,, Т — гендер символы: О —хромосоманың донордан рециниентке беріле бастайтьщ учаскесі.
Жыныстыц фактор Ғ+-тщ хромосомалық еместігі, оның ДНК-дан тұратыны және онда нуклеотидтердің шамамен 105 жұбы болатыны бірқатар эксперименттер арқылы дәлелденген.
Қазіргі уақытта ДНК-ның автономиялы репликацияға қабі-летті, бөліну кезінде жас клеткаларға берілетін және белгілі бір белгілерді анықтайтын айтарлықтай көп, хромосомадан тыс молекулалары белгілі. Олар плазмидтер деп аталады. /7+-фак-тор—ішек таяқшасының жақсы зерттелген плазмидтерінің бірі. Ғх —■ плазмидтердің өзгерген басқаша туріне жатады. Ол бол-ған жағдайда донор клетка бұл плазмидті реципиентке беріп қана қоймастан, сондай-ақ хромосомалық гендерді беруге де қабілетті болады.
Ішек таяқшасының басқа штаммаларының ішінен рекомби-нацияның жоғары жиілікте болуын анықтайтын және Щг таң-басымен белгіленген тағы бір жыныстық тип анықталды. Ғ~ X Н^г шағылысуы рекомбинанттардың өте жоғары процен-тін береді: 10 бастапқы клеткаға 1. Сонымен қатар, Ғ+ клетка-лармен шағылыстырудан өзгеше, Ғ~ «аналық» клеткалары Я/Ү-дің қасиеттерін ете сирек жағдайларда қабылдайды. Бұл жағдайда жыныстық фактор басқа гендермен тіркесе отырып берілетінін және бактериялық хромосомада белгілі бір локусты алып жататынын генетикалық талдау көрсетті. Хромосома Ғ~ клеткасына ауысқанда ол әрқашан да соңғы болып шығады (78-сурет, 2).
У. Хейс Щг штаммалары Ғ+ штаммаларынан шығатынын және бұл өзгеріс фактордың жойылуымен байланысты еместі-гін көрсетті, өйткені кері мутациялануда Н}г-+Ғ+ жыныстық фактордың донорлық қасиеті қалпына келеді. Сонымен, егер клеткада Ғ-фактор болса, ол өзін екі түрлі, яғни цитоплазмалық бөлшек ретінде (Ғ+ клеткаларында) немесе хромосоманың локусы ретінде (Н[г клеткаларында) ұстауы мүмкін.
Бактериялық клетканың хромосомасына ене алатын плаз-мидтерді эписомалар деп атайды. Плазмидтер геномы оларды жеткізетін бактериялардың қасиеттерін анықтайды. Қасиеттері әр түрлі болуы мүмкін, бірақ плазмидтер көп ретте бактерия-лардың ауыр металдардың түздарына, антибиотиктерге және басқа агенттерге төзімділігін анықтайтын ферменттерді жасап шығаруға жауапты болады. Кейде олар бактериялардың ауру туғызғыштығын да анықтайды. Дәрі-дәрмектерге резистентті-лікті анықтайтын плазмидтерді і? таңбасымен белгілейді. Олар ішек таяқшасының бір мезгілде, мысалы, 10-нан астам антибио-тиктерге төзімділігін анықтай алады. Жапонияда дизентерия аурулары терапиясында антибиотиктерді қолданудан кейін сипатталған ғажайып құбылысты олардың дәл осы қасиеттерімен түсіндіруге болады. Аурулар саны кенет кеміді де, содан соң қайтадан тез өсе бастады. Осы уақытта көптік төзімділік, яғни /?-плазмидтері бар бактериялардың кездесу жиілігі де артты. Резистенттілікті қамтамасыз ететін механизмплазмидтердің антибиотиктерді бүзатын ғіемесе инактивтендіретін ферменг синтезін детерминациалау иемесе клетканың өткізгіштігін оған антибиотик енбейтіндей етіп өзгерту қабілетімен анықталадьь Плазмидтер молекулалық массасы 1-108 Д-дан аспайтын цир-кулярлық формасы бар ДНК молекулаларынан түрады. Плаз-мид реципиент клеткасына бір созынды ДНҚ түрінде беріледі де,, онда клетка мембранасына бекітілген күйінде екі еселенеді. Осыдан кейін оның жабысқақ үштары шеңбер жасай жабысады да, едәуір күрделі спиральданған қүрылымға айналады. Әр тур-лі плазмидтердің репликациясы хромосомалық ДНҚ репликациясы секілді тәуелсіз реттеледі: бұл белгілі бір жағдайларда клеткада белгілі бір плазмидтердің ғана талғамалы реплика-циялануына әкеп соғады.
4. РНК — ТҰҚЫМ ҚУАЛАУ ИНФОРМАЦИЯСЫН САҚТАУШЫ
Микроорганизмдер генетикасын зерттеу көптеген жаңа қү-былыстарды ашуға мүмкіндік берді, оның ішінде Місоііапз туысына жататын өсімдіктерді зақымдайтын темекі мозарікасы вирусында түқым қуалау информациясын РНҚ молекуласы жеткізетіні дәлелденді.
Бұл былайша дәлелденді. Вирус спираль түрінде ширатылған РНК молекуласынан басқа, сондай-ақ белокты қабықша-дан түрады. Егер РНҚ-дан белокты бөліп алатын болсақ, РНК-дан өзгеше, оның темекі жапырағына тигізгенде осы ауруға тән некроздық дақтар туғызатын жүкпалы қасиеті болмайды. Бел-гілі бір жағдайларда РНК мен тіпті әр түрлі вирустардан алынған белокты пробиркада араластыра отырып, вирус бәл-шектерін қалпына келтіруге болатындығы көрсетілді. Темекі мозаикасы вирусының 2 штаммасы алынып, РНҚ мен белокқа бөлінді. Осының нәтижесінде 4 компонент алынды, осыдан кейін олар мүмкін болатын барлық тәртіппен араластырылды да, 4 түрлі—2 бастапқы және 2 жаңа «будандық» бөлшектер алын-ды, оларда бір штамманың РНК-сы, екінші штамманьщ белогы болды. Темекі мозаикасы вирусының «будандық» бөлшектері ■оларды өсімдіктерге жұқтырғаннан кейін жаңа вирустық бөл-шектер репродукциялады. Бір қызығы сол, бұл синтездік бу-дандардың РНК-сы тек өз белогын ғана, яғни одан РНҚ бөлін-ген штамманың белогын синтездеген. Жұқтырудан кейін темекі мозаика вирусының бастапқы штаммалары ғана қалпына кел-ген, ал «будандық» формалар өзін өндіре алмады.
Бүл мәліметтерді, сірә, темекі мозаика вирусында белоктың құрылымын анықтайтын тұқым қуалау информациясы РНК-да болатындығымен ғана түсіндіруге болар.
Жануарлар мен адамның клеткаларында паразиттіктіршілік «тетін вирустардың ішінде ДНК-сы емес, РНҚ-сы бар формалар белгілі (мысалы, полиомелитті, энцефалитті және т. с. с. туғы-затын вирустар). РНК-сы бар бактериофагтар да сипатталған.Сонымен, жаңа қүбылыстарды (трансформация, трансдук-ция, плазмидтер мен эписомалардың болуы) ашу микроорга-Бизмдерді зерттеуде түқым қуалаушылықтың материалдық не-гіздері туралы біздің түсініктерімізді едәуір кеңейтті және ДНК-ның клеткадағы құрылысына (жекелеген молекулалар немесе хромосомалар) және шоғырлануына (ядро немесе цито-плазма) қарамастан, клеткадағы түқым қуалау информациясын сақтау ролін ДНК атқаратынын сенімді түрде көрсетті. Кейбір вирустар мен фагтарда түқым қуалау информациясын сақтау-шы РНҚ болуы мүмкін.
16-тарау. ГЕННІҢ ТАБИҒАТЫ
Тұқым қуалаушылықтың дискретті бірлігі болатынын Мен-дель айқын жорамалдаған. Морган геннің хромосомалармен байланысты екенін көрсетті.
Рекомбинацияның, мутацияланудың және фуғікцияның бір-лігі ретіндегі ген туралы генетиканың ерте кезеңіндегі түсінік оның бұдан былайғы дамуыиа қолайлы әсер етті. Кейінірек ген-нің бүған дейін есептеліп келгеніне қарағанда, түқым қуалаутшылық пен өзгергіштіктің едәуір күрделі бірлігі екені анықтал-ды. Бүл тарау ген туралы осы кездегі түсініктерді талдауға арналады.
1. АЛЛЕЛИЗМ ЖӘНЕ АЛЛЕЛИЗМ КРИТЕРИЙІ
Бір геннің әр түрлі күйін аллельдер деп атайды. Мутацияла-ну нәтижесінде ген екіден артық (көптік аллелизм қүбылысы, 13-тарауды қараңыз) күйде бола алатыны белгілі.
Мынадай сұрақтың тууы заңды нәрсе: белгілі бір белгінін. көрінісін өзгертетін, бір-бірінен тәуелсіз пайда болған қандай да екі мутацияның аллельді немесе аллельсіз екенін, яғни олардың бір генде немесе әр түрлі генде пайда болғандығын іс жү-зінде қалай анықтауға болады? Аллелизмнін, критерийі қандай? Бүл сүрақтарға алғаш рет Т. Морган жауап берді. Ол алле-лизмнің екі критерийін ұсынды: функционалдық (немесе комп-лементарлық) және рекомбинациялық.
Функционалдьщ критерийі әр түрлі гендердің өзгерісі бар екі мутантты шағылыстырғанда бірінші үрпақтың буданы — әрбір геннің қалыпты аллельдерінің басым болуына байланыс-ты жабайы фенотипі бар дигетерозигота пайда болатынына негізделеді. Мұндай жағдайда зерттелетін мутациялардың қа-лыпты аллельдері бір-біріне комплементарлы деп есептеу қа-былданған. Сонымен қатар, егер шағылыстырылатын мутанттарда бір геннің аллельдері болса, компаундта жабайы тип пайда болмайды. Мысалы, екі мутантты — ақ және сарғыш күзен- ді шағылыстырғанда барлық будандар қоңыр реңді, яғни жа- байы фенотипті болады. Ақ күзенді басқа мутанттық форма —■ платина реңдес күзенмен шағылыстырғанда да барлық будан-дар платина реңдес, яғни мутантты фенотипті болады. Демек, бірінші жағдайда комплементарлық, яғни аллель еместік; ал екінші жағдайда — комплементарлықтың жоқтығы, яғни ал-лельдік байқалады. Бүл жайында «Генетиканың практикалық сабақтарына басшылықта» толығырақ жазылған.
Әр түрлі гендердеғі мутациялар ғана өзара рекомбинациялана алады деген түсінік рекомбинацияльщ тестің негізіне алынды. Морган мектебінің зерттеушілері, функционалдық (гетеро-зигота — мутантты фенотип) және рекомбинациялық (рекомби-нация жоқ) критерийлер сақталған жағдайларда мутациялар-ды аллельді деп есептеді, Геннің қүрылымы туралы түсініктің өзгеруіне байланысты аллелизмнің критерийі де анықталды, ол туралы алда айтылатын болады.
2. ГЕННІҢ ҚҰРЫЛЫМЫ
Сатылы аллелизм. Бүрын ойлағанға қарағанда геннің зор функционалдық тұрақсыздығын білдіретін көптік аллелизм қүбылысының табылуы (13-тарауды қараңыз) геннің күрделілі-гін көрсететін алғашқы дәлелдемелердің бірі болды.
1929—1930 жылдары біздің елімізде А. С. Серебровский мен оның жас қызметкерлері — Н. П. Дубининнің, Б. Н. Сидоровтың және т. б. жүмыстарында геннің функционалдық күрделілігі түңғыш рет эксперимент арқылы көрсетілді. Авторлар дрозофиладан жыныстық хромосоманың нольдік нүктесіне шо-ғырланған локустың зсиіе көптік аллельдер сериясын зерттеді. Бұл локустың 5С1, 5С3 және басқа мутациялары шыбын дене-сіндегі әр түрлі қылшықтардын, редукциялануына себепші бола-
ды. ==^5 және ==|= особьтарын шағылыстырғанда екі генге жатқызылуы мүмкін екендігі көрсетілді, Мутациялар жүрген барлық 300 нүкте көрсетілді, мүның ішінде кейбір нүктелерде олар өте жиі болған (мысалы, зерттелген 149 мутация-ның 123-1 бір нүктеде өткен). Мұндай нүктелер немесе-орындар «негізгі дақтар» деген атаққа ие болды. Ген ішінде мұндай нүктелердің орналасуы кездейсоқ қүбылыс емес.
Әр түрлі объектілердің «негізгі нүктелерінің» ортақ белгіле-рі болатынын, яғни бірдей нуклеотидтердің ДНК тізбегіне, мы-салы, А Ц А А А А А А Г, яғни 6 адениннің қатар шоғырланаты-нын зерттеулер көрсетті. Бұл мәліметтер бірдей нуклеотидтер-дің ДНК-де орналасу реті осы локустардың жоғары жиілікте мутациялануының себебі болуы мүмкін деп жорамалдауға мүм-кіндік береді.
Геннің молекулалық құрылысы. Геннің шамасын, яғни онда-ғы нуклеотидтер жүбының санын және оның минимальды моле-кулалық массасын шамамен есептеп шығаруға болады. Қазіргі уақытта Г4 фагының 50-ден астам гені белгілі. Объектіні зерттеу барысында гендер саны арта түсері сөзсіз. Ол екі еселенеді деп жорамалдауға болады. Бүл фаг ДНК-ның молекулалық массасы 120Х^06-ға тең. Демек, бір геннің молекулалық массасы 1 X 106 шамасында болады. Нуклеотидтердің бір жүбының молекулалық массасы 600-ден сәл асатындықтан, бүл ген орта есеппен алғанда нуклеотидтердің сызықтық орналасқан 1500 жұбынан түратындығын білдіреді. Жуықтатылып алынған бүл есептеулер геннің атқаратын қызметі мен оның мөлшері (500— 6000 нуклеотид) туралы осы кездегі түсініктермен жақсы үйле-седі. С. Бензер Г4 фағының г II ауданы картасының генетикалық және физикалық (молекулалық) масштабтарын салыстырып, Т4 фагының рекомбинация бірлігіне ең артық дегенде нуклео-тидтердің екі жүбы келетінін, ал жекелеген нуклеотидтер мута-циялар бірлігі болуы тиістігін есептеп шығарды.
Аллель аралық комплементация. 50-жылдардың аяғында функционалдық тест аллелизмнің абсолюттік критерийі бола алмайтыны көрсетілді. Бір геннің кейбір мутациялары транс жағдайда комплементарлы болуы мүмкін, яғни гетерозиготалы күйдегі (а1/а2) мутанттық аллельдер жабайы немесе оған жа-қын типтің фенотипін анықтауы мүмкін. Бүл құбылыс ген іші-лік немесе аллелъ аралық комплементация деп аталды. Аллель аралық комплементация механизмі әлі жеткілікті зерттелмеген. Алайда, бұл жағдайда жабайы тип белгІлерінің пайда болуы цитоплазмада белоктар — екі мутанттық аллельдер өнімдері-нің арасындағы езара әсер ету есебінен өтетіндігі көрсетілді. Мүндай өзара әсер ету молекулалары үқсас суббірліктерден тұратын ферменттер (белоктар) үшін ғана мүмкін болады. Бүл жағдайда гетерозиготалы особьта ұқсас, бірақ әр түрлі кемші-ліктері (мутанттық) бар суббірліктер ферменттің бір молеку-ласына бірігеді және осы кезде езара түзету процесі жүреді: белоктың ферменттік активтілігі қалпына келеді. Бұл эффект ашытқы, нейроспора және басқа микроорганизмдер үшін, содай-ақ дрозофила үшін де көрсетілген. Ол өзін-өзі іп юііго өнді-реді.
* * *
Сонымен, геннің құрылысы күрделі екендігі айқын, ол крос-синговерде бөлінетін бірлік болып табылады да, мутациялану бірлігі болып саналмайды. Алайда белок молекуласын оны маманданған қызметінен айырмайынша бөліктерге бөлуге болмайтыиы секілді, тұқым қуалау информациясының бірлігі ре-тінде ген де атқаратын қызметі жағынан бөлінбеген күйінде қалады. Аллелизмнің қолданылыр жүрген функционалдық және рекомбинациялық критерийлері' салыстырмалы.
17-тарау. ГЕНЕТИКАЛЫҚ КОД
ДНҚ геннің ерекше компоненті болып табылатындығын дә-лелдейтін сенімді мәліметтер алынған соң мынадай мәселелер алға койылды: ДНҚ-ның тенетикалық қүрылымы (тұқым қуа-лау информациясын жеткізущі ретінде гениің алғашқы функ-циясы) қағідай? ДНҚ организмнің онтогенездегі белгілері мен қасиеттерін (екінші функциясы) қалай анықтайды?
Мендель — Морганның ген ■— белгі постулаты генетикалық зерттеулердің негізгі өзегі болды. Гендердің көпшілігі ерекше белок-ферменттер синтезін бақылайтындьғғы көптеген микроор-ганизмдерден көрсетілді: гендер мутациясы ең алдымен тиісті ферменттер активтілігінің езгеруінен немесе жойылуынан бай-қалады. 13-тарауда айтылғанындай, адамда фенилкетонурия және т. б. осы секілді аурулардың пайда болуы жекелеген ген-дердің мутациялануы салдарынан ферменттік қызметтің бүзы-луымен байланысты болады. Бұл фактілер Г. Бидл мен Е. Та-тумға «бір ген — бір фермент» қағидасын үсынуға мүмкіндік берді. Қазіргі уақытта бұл қағиданы әрбір белок үшін оның қүрылымы мен активтілігін бақылап отыратын ген болады деп түсіну керек.
Алынған жаңа фактілердің әсерімен бүл қағида одан әрі нақтыланып, анықтала түсті. Адам гемоглобинінің терт — екі а және екі |3 тізбектен түратын молекуласындағы әрбір тізбек генетикалық жолмен жекелей бақыланатыны анықталды. Осы және осыған үқсас басқа да фактілер «бір ген — бір полипеп-тидтік тізбек» деген неғұрлым дәл постулатты енгізуге мүмкін-дік берді. Ал енді полипептидтік тізбекте, дәлірек айтқанда генетикалық түрғыдан не бақыланатынын анықтап көрейік. Адам қанының тұқым қуалайтын зілді ауруы — орақша-клет-калы анемия ауруы кезінде — мутацияның нәтижесінде гемо-глобиннің қасиеті езгереді. Бүл жағдайда 574 амин қышқылы-ның қалдығынан тұратын гемоглобин молекуласында бір ғана алмасу өтеді: глутамин қышқылын валин алмастырады. Осыдан келіп, ген полипептидтік тізбектің амин қышқылды қүрамын, демек, олар түзетін белоктарды бақылайды деген қорытынды жасауға болады.
1. ДНК-ның ГЕНЕТИКАЛЫҚ ҚҰРЫЛЫСЫ
Жүйелілік болжамы. ДНҚ-да информация қалай жазылған? Бұл сұраққа жауап беру үшін, бір жағынан, ДНК молекуласы-ның (2-тарауды қара) екінші жағынан, белоктардың құрылы-сын еске түсіру керек. ДНК-нің құрамына оның ерекшелігін айқындайтын 4 негіз (аденин және гуанин — пуринді, тимин жәие цитозин — пиримидинді негіздер) кіреді. Белоктардын, құрамына 20 амин қышкылы кіреді. Міне осылар, полипептид-тік тізбектегі осылардың орналасу жүйелілігі белок молекула-ларының алғашқы күрылымыи ғана емес, сондай-ақ оның төр-тінші құрылымына дейінгі бүкіл кейінгі қүрылымын, демек, белоктардың бүкіл алуан түрлілігі мен ерекшелігін анықтайды. Сонымен, нуклеотидтердің ДНК-да орналасу жүйелілігінде амин қышқылдарының белок молекуласында орналасу жүйёлі-лігі туралы тұқым қуалау информациясы жазылуы мүмкін деп жорамалдауға болады.
Бүл қағиданы жүйелілік болжамы түрінде Ф. Крик дамыт-ты. «Жүйелілік болжамының» әділдігі, атап айтқанда, Ч. Янов-ский мен оның қызметкерлерінің жүмыстары арқылы дәлелден-ді. Авторлар Езсһегісһіа соіі-дегі триптофансинтетаза фермен-тін бақылайтын локуетың нәзік қүрылымын зерттеп, ДНК молекуласындағы нуклеотидтер жүйелілігінің өзгеруі болып са-налатын тоғыз мутацияны тапты. Мутанттық форманың белок-ферментіндегі амин қышқылдарының орналасу 'жүйелілігін зерттеу оның ДНК-дегі нуклеотидтер жүйелілігінің өзгеруіне параллельді өзгеретінін көрсетті, яғни геннің және ол бақылайтын белоктьң колинеарлығы дәлелденді.
Бірақ тұқым қуалау информациясы ДНҚ-да қалай жазыл-ған, қалай шифрланған? Генетикалық информацияның гендер-де кодпен 1 жазылғандығы туралы идеяны алғаш рет 1954 жы-лы Д. Гамов айтқан.
Генетикалық кодтың негізгі белгілері. Генетиктердің, био-химиктердің, цитологтар мен басқа да мамандардын, күш салуы арқасында қазірғі уақытта генетикалық кодтың негізгі белгіле-рі бөлгілі болды.
Код ең алдымен триплетті болып табылады, яғни әрбір амин қышқылын үш негіз (кодон) кодпен береді. Төрт негізден 3-тен әр түрлі 64 комбинация жасауға болатынын қарапайым есептеу көрсетеді, бүл 20 амин қышқылын кодпен жазуға толық жетжі-лікті^Бірақ, теориялық есептеулерден басқа, Ф. Крик пен оиың қызметкерлері ішек таяқшасын зақымдайтын Г4 фагының гІІ мутациясын зерттеуде кодтың триплетті екендігіне тікелей эксрерименттік дәлелдемелер алды.
Шуклеотидтердің үш жүбының ұштасуын авс деп белгілейік. Бүдан соң мүндай триплеттер ДНК молекуласының кесіндісінде бір жүйелілікпен қайталанып отырады деп есептейік: Бір объектіні басқа объектілердің жәрдемімен бейнелеуді кибернетика-да кодпен жазу деп атайды.
авс авс авс \ авс..., ^ Енді, мутация жүрді, мысалы, артық нуклеотид а қосылды (стрелкамен көрсетілген) деп есептейік, егер код солдан оңға қарай, яғни әрқашан да бір бағытта үштікпен оқылатын болса, онда («тексті») триплеттерді оқудың бүкіл бұдан былайғы тәр-тібі бүзылады:
авсаавсавсавс-^-авс аав сав сав... Осыдан кейін бастапқы мутациядан сәл оңға таман белгілі бір негіздің, мысалы, с-тің (стрелкамен көрсетілген) түсіп қалға-нын білдіретін жаңа мутация пайда болды дейік: авсаавсававс-^авс аав сав авс... Бүл жағдайда екі мутацияның арасындағы учаскеде ғана (стрелкалармен көрсетілген) «текст» бүзылады, ал учаскенің қалған бүкіл үзына бойына бұрынғы оқу тәртібі сақталады. Қү-рамына бірнеше өзгерген триплеттер кіретін геннің осы бөлігі қандай да бір жауаптылығы шамалы функцияны атқаратын болса, онда белгілі бір шағын учаскеде кодтың бұзылуы ақырғы нәтижеге айтарлықтай әсер етпейді деп түсінуге болады. Мүндай екі өзгерісі бар особьтарда жабайы типке жуықтайтын федртип болады, бүл жағдай эксперимент арқылы көрсетілді.
? Егер код шын мәнінде триплетті болса, негіздердің екі қосиасының (немесе (екі түсіп қалуының) комбинациясы/алдыңғы жағдайдан өзгеш^, әрқашан да мутантты фенотип беруі тиіс. Бүл да эксперименттік жолмен дәлелденген. Егер жабайы типтің Г4 фагында нуклеотидтердің (2-жол) және олар. арқылы анықталатын амин қышқылдарының (1-жол) жүйелілігі мына-дай:
Лиз Сер Про Сер Лей Асп Ала ААА АГУ ЦЦА УЦА ЦУУ ААУ ГЦУ болса, Г және У (жақшамен белгіленген) екі нуклеотидін қос-қанда мынадай жүйелілік алынады:
ААА АГУ ГУЦ ЦАУ ЦАЦ УУА АУГ Лиз Сер Вал Гис Гис Лей Мет. Осыған орай белоктың (төменгі жол) амин қышқылдық қүра-мы да өзгереді, бүл жағдай мутация ретінде тіркелген.
Сонымен, шағылыстыру нәтижелері барлық жағдайларда алдын ала болжанған нәтижелерге сай келді, бұл кодтың трип-леттігі туралы болжамның дәлелі болды.
Алдыңғы пікірлерден байқалғанындай, код бірін-бірі жаба алмайды, яғни белгілі бір нуклеотид көршілес екі триплеттін. құрамына бір мезгілде ене алмайды. Бүл Криктің біз айтып әт-кен жүмыстарында эксперименттік жолмен дәлелдендьі Алайда бүл қағиданы дәлелдейтін тікелей эксперименттер де жүргізіл-ді. Азотты қышқылдың жәрдемімен темекі мозаика вирусынан бір ғана негіздің (цитозин урацилге) алмасуына байланысты болатын мутация алынды. Осы вирус синтездейтін қабықшаның белогында бір ғана амин қышқылының өзгерісі көрсетілді. Адамның мутанттық гемоглобинін талдағанда да осындай нә-тижелер алынды.
/Келтірілген есептеулерге сүйенсек (20 амии қышқылына 64 кодон) код «өзгерген» болуы тиіс, яғни белгілі бір амин қыш-қылын бірнеше кодон анықтауы тиіс. Бұл есептеулер сондай-ақ «мәнсіз» триплеттердің, яғни амин қышқылдарының бірде-біреуін кодпен белгілемейтін /гриплеттердің болатынын болжауға мүмкіндік береді. Бұл болжамдар эксперименттік жолмен дә-лелденді, олар туралы кейінірек айтылады.
Кодтың өзіне тән сипаты: оны оңу арқашан да белгілі бір пункттен басталады. Г4 мутантты фагының рекомбинациясын зерттеу арқылы Крик бұған да эксперименттік дәлел тапты. ^
Негіздер қосылатын және түсіп қалатын эксперименттерде код үтірсіз оқылатыны, яғни бір нуклеотид түсіп қалатын бол-са„ оқу кезінде оның орнын кәршілес кодонның жақын орналас-қан нуклеотиді алатыны, осының арқасыида бүкіл оқу тәртібі өзгеретіні керсетілді.
Соңғы жылдары ДНК молекуласын нукл.еотидтері белгілі бір жүйелілікпен орналасқан жерлерден (мысалы, аденин мен гуанин аралығынан) езіне тән ерекшелікпен қиып түсетін фер- мент-рестриктазаны пайдаланғанда фХ174 фагында бірін-бірі жабатын гендер болатындығы жәнінде жаңа мәліметтер алын-ды. Бүл осы фаг ДНК-ның шамамен алғанда 5400 нуклеотидтен түратын молекуласында гендер былай орналасқан: геннің бас-тапқы үш нуклеотиді басқалары үшін аралық болып табыла-тындай болып орналасқан. Мүнда осы гендер нуклеотидтерінің триплеттері сәйкес келмейді. Мысалы,
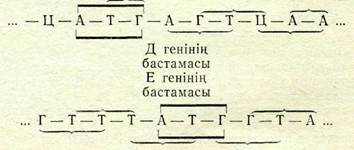
Алайда, бірін-бірі жабатын гендер табиғатта қаншалықты жиі кездеседі және олардың ролі қандай деген мәселелер әлі де шешілмеген куйде қалуда.
2. БЕЛОК СИНТЕЗІН ГЕНЕТИКАЛЫҚ БАҚЫЛАУ
Белок синтезінің механизмі. Код туралы мәліметтер алынған соң белок синтезінде ДНК қандай роль атқарады деген сүрақ туды.
■ Ген — ДНК учаскесі — белгілі бір жүйелілікпен орналасқан
 |
| РЕПЛМКАЦИЯ ар |
| Актидтендіруші "(рермент |
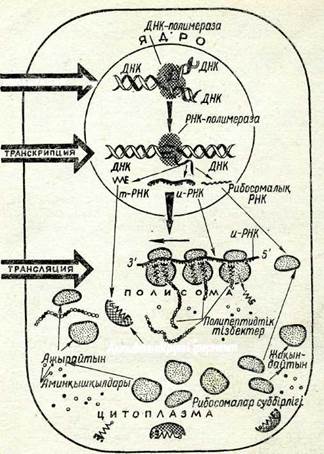
80-сурет. Клетка-дағы белок синте-зінің схемасы.
Амин қышқылдары бар полипептидтік тізбектерді синтездеуге арналған информация болады, алайда ген қалыпты қызмет атқару үшін оған жол ашатын және геннің бастамасын анықтай-тын нуклеотидтердің белгілі бір жүйелілігіне — промоторға жә-не одан кейін келетін, геннің аяқт.алуын білдіретін жүйелілік — терминаторға ие болуы тиіс. Гендер арасында, әдетте, әзірше ролі анықтала қоймаған жүйелілік бар. Ген ДНК-сы белок син-тезіне тікелей қатыспайды. ДНК геннің алғашқы өнімі — информациялық РНК (и-РНК) молекуласының синтезі үшін ма-трица болыи қана қызмет атқарады.і,
Қазіргі уақытта ДНК спиралінің тізбекшелері бір мағыналы емес таңбалы деп есептелінеді: олардың бір тізбекшесінде поли-пептид қүрылымы шифрланған — оның мағыналық мәні бар, ал оған комплементарлы — екіншісі мағынасы барға қарсы тізбек болып табылады. и-РНК-ның бір қабат тізбегінің синтезі (транскрипциясы) үшін мағыналық тізбекше ғана матрица бола алады (80-сурет),
ДНК-ның мағыналық тізбекшесінің кодын и-РНҚ-ға көші-ру РНК-полимераза (транскриптаза) ферментінің бақылауымен жүзеге асады, мұнда и-РНҚ ДНҚ-ның мағыналық тізбекшесі-не комплементарльиіБұл егер бір тізбектегі жүйелілік белгілі болса, екінші тізбектегі жүйелілікті автоматты түрде шығаруға болады деген сөз. Мәселен, ДНК-дағы валин синтезін бақылау-шы кодон — ЦАА болса, и-РНҚ-да ол ГУУ болады. Клеткада и-РНҚ молекуласыиың тіршілік ету уақыты әр түрлі. Мысалы, бактерияда ол бірнеше минут, бидайдың қүрғақ тұқымының клеткаларында — бірнеше ай, жануарлар ретикулоциттерінде и-РНҚ гемоглобиннің а және р-тізбекшелерін синтездеу үшін клетканың бүкіл әмірі бойында тіршілік етеді. Синтездеуден кейін и-РНК молекуласы ДНК-дан бәлінеді де, рибосомалар-мен жанасады. Оның үстіне жоғары сатыдағы организмдер мен кейбір вирустарда и-РНК молекуласының (5'-шеті деп аталатын) бір шетінде бір немесе екі метильденген негіз болады, көп ретте бұл жетінші жағдайда қүрамында СН3 тобы бар гуанин болады. Рибосомалар белок синтезі үшін бастапқы ретінде осы үшін тандап алады.^и-РНК мен бірнеше рибосоманың (2-ден 15-ке дейін) комплексі полирибосома немесе полисома деп ата-лады. Белок синтезі (трансляция) осы полисомаларда өтеді. Ри-босомалар үлкен полимерлі рибосомалық РНК (р-РНҚ) мен белоктан түратын құрылым болып табылады. Соңғы уақытта көрсетілгеніндей, белок синтезінің инициациясына р-РНК-нын, тікелей қатысы бар. Рибосома клеткада әр түрлі күйде бола алады: екі суббірлік түрінде (седиментация цифры 30—405 кіші және седиментация цифры 50—605 үлкен суббірліктер) және аталған бөліктерден түратын бірыңғай қүрылым түрінде. Оның күйі атқаратын қызметіне байланысты. Егер рибосома дәл осы мезгілде белок синтезіне қатыспайтын болса, онда ол клеткада жеке суббірлік түрінде кездеседі. Синтез кезеңінде кіші суббір-лікке и-РНҚ молекуласы бекінеді. Бүдан соң оған ірі суббірлік қосылады да, рибосома әзіне тән пішінге ие болады (80-сурет).
Әрбір рибосома и-РНК молекуласын бойлай 5'-шетіңен 3'-шетіне қарай қозғалады, бұл уақытта полипептидтік тізбекше өсе береді. Тізбекшенің үзындығы рибосоманың и-РНК моле-куласының басынан бастап өткен қашықтығына пропорционал-ды болады. Мұндай белок фрагментіндегі амин қышқылдары-ның саны рибосома оқып үлгірген триплеттердің санына тең бо-лады.
и-РҢҚ триплеттеріне (кодондарына) сәйкес рибосомаларда амин қышқылдары қалдықтарының тізілуі РЬіК молекулалары-ның тағы бір түрінің, атап айтқанда транспорттық РНҚ (т-РНҚ)-ның.жәрдемімен жүзеге асады. т-РНК төмен полимерлі болады, оның жекелеген молекулалары 70—80 нуклеотидтерден түрады. Негіздердің өзара комплементарлы әсер етуінің арқасында бүл молекулада сутектік байланыстардың көпшілігі түй-ықталған. т-РНК қүрылысы модельдерініц біріне сәйкес оның молекуласының пішіні беде жапырағының пішініне ұқсайды және оған амин қышқылы қосылатын деп жорамалданады. т-РЫК молекулаларының қарама-қарсы бөлігінде антикодон деп атала-тын қосарланбаған 3 нуклеотид болады. т-РНК антикодоны-ның триплеті сутектік байланыс түзе отырып, рибосомадағы и-РНҚ кодонының триплетімен әрекеттесе алады. т-РНҚ молекулаларының әрбір класы бір ғана амин қышқылымен қо-сыла алады: демек, әр түрлі т-РНК. молекулаларының мөлшері жиырмадан кем болмауы тиіс. Биохимиктердің мәліметі бойын-ша олар — 60. Қазіргі кезде олардың кейбіреуінің химиялық құ-рылымы анықталған. Совет ғалымы академик А. А. Баев алғаш рет валинді т-РНК-ның құрылымын анықтады.
т-РНК-ның амин қышқылдарымен әрекеттесуі үшін амин қышқылдары АТФ-ның қатысуында амино-ацил-т-РНК ерекше ферменттерімен активтенуі қажет. Сонымен қатар, т-РНҚ мен активтелген амин қышқылдарының өзара әрекеттесуін активтен-діруші аса ерекше ферменттер т-РНК-лигазалар (немесе кодазалар) бақылап отырадьі^і-РНК-ның амин қышқылдарымен әрекеттесуін кейде белок синтезінде жүмыс істейтін клеткадағы екінші кодтық система деп атайды.
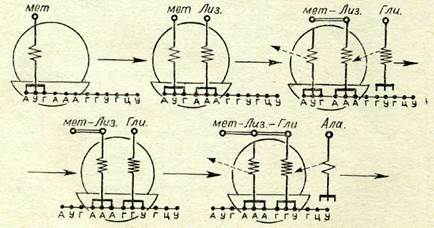 |
| 81-сурет. Трансляцияның жүйелі кезеңдерінің схемасы (түсінігі тексте берілген). |
. т-РНК молекуласын рибосоманың ірі суббірлігі белгілей-ді, бір оның бүған арнаулы екі орнй болады. Белок синтезін инициациялаушы и-РНК кодоны — АУГ құрамында метионин бар т-РНК-ны ажырата біледі. Бүдан соң осы рибосомаға ке-лесі кодонға сәйкес екінші т-РНҚ бекінеді. Амин қышкылда-рының арасында пептидтік байланыстар пайда болады (бұл про-Цеске де ферменттер қатысады). Өзіне түскен салмақтан арыл-ған бірінші т-РНҚ рибосомадан шығады. Келесі амин қыш-қылына пептидтік тізбекті қосып алған соң екінші т-РНК да рибосомадан шығады. и-РНК триплеттерінің жүйелілігіне сәй-кес полипептидтік тізбектің есуі осылайша өтеді (81-сурет).
Генетикалық информацияны оқудың дәлдігі рибосома арқы-лы қандай кодондар өтетіңдігіне ғана емес, сондай-ақ едәуір дә-режеде рибосоманың өзінің күйіне де байланысты. Стрептоми-цин антибиотигінің молекулаларын қосу арқылы рибосоманың құрылымын өзгертсе немесе қоршаған ортадағы М§++ иондары-ның конценлрациясын арттырса болғаны, и-РНК-да жазылған генетикалық код бұрыс оқыла бастайды: рибосома қате «оқи» бастайды, полипептидтік тізбекке көп ретте «басқа» амин қыш-қылдарын енгізеді. Стрептомициннің бактериялық клеткаларға инактивациялаушы әсері, міне осы жағдайға байланысты. Қа-лыпты жағдайда белок синтезі қатесіз әрі үлкен жылдамдықпен өтеді: 1 секундта 2 амин қышқылы түзіледі. Белоктыц 150 амин қышқылынан түратын молекуласы бір полисомада шамамен 1,5 минутта синтезделеді.
Қатерлі ісік (тауықта Раус саркомасын, тышқанда лейкоза-ны) туғызатын вирустарды зерттеуде Г. Темин мен Д. Балткмор 1970 жылы жаңа фермент — РНҚ-ға тәуелді ДНҚ-полимераза, немесе «кері транскриптаза» иемесе ревертаза ферментін ашты. Бұл фермент генетикалық информацияны әдеттен тыс бағыт-та — РНК-дан ДНҚ-ға қарай тасымалдауды жүзеге асырады. Бүл процесс вирус РНҚ-ында матрицадағыдай ДНК синтезде-ліп, соңынан клетка-иесінің хромосомасына ене алатын жағдай-да жүзеге асады. Кері транскрипцияның болатындығын 1960 жылы СССР-де С. М. Гершензон тапқан болатыд/ Бірақ бұл жа-ңалық ферментті бөліп алған соң ғана тиісінше бағалана баста-ды. 1971 жылы тышқан мен адамның вирус жүқпаған клеткала-рынан осындай фермент табылды. Олардың шығу тегі және қа-лыпты клеткалардағы түзілуі мен атқаратын қызметі әзірше белгісіз, ол бүл жағдайда да онкогендік виру.стармен байланыс-ты болар деп жорамалданады. Бұл жаңалық тіршілік процесте-рін түсіну үшін, сондай-ақ рак проблемасын шешу үшін көпте-ген пайда келтіретін болады (29-тарауды қара).
Белок синтезін генетикалық бақылау. Белок синтезі механиз-мінің негізгі кезеңдерін түсіну осы процесті іп ^ііго модельдеуге мүмкіндік берді. Биохимиядағы екі жаңалық: белок синтезі жү-зеге аса алатын клеткасыз системаны алу және жасанды поли-рибонуклеотидтерді ферменттік синтездеу осылай етуге мүмкін-дік туғызды.
1961 жылы М. Ниренберг пен Г. Маттэй қүрамында рибосо-ма, амин қышқылдарының толық жиынтығы және кейбір фер-менттер бар клеткасыз системаға енгізілген, жасанды жолмен синтезделген, белгілі бір қүрамды полирибонуклеотидтер оған и-РНК ретінде әсер ете алатындығын кәрсетті. Жасанды жол-мен синтезделген, қүрамына тек қана урацил (полиуридил қыш-қылы, поли-У) енген информациялық РНК-ны Езсһегісһіа соіі- ден алынған клеткасыз системаға орналастыру арқылы белгілі бір қүрамды белокты синтездеуді қамтамасыз етуге мүмкіндік туды. Синтезделген полипептидті талдағанда ортада барлық амин қышқылдарының болуына қарамастан, оның құрамына бір ғана амин қышқылы, атап айтқанда, фенилаланин кіргені анық-талды, яғни полиуридил қышқылы полифенилаланин синтезде-луіне жағдай жасаған. Сонымен, генетикалық кодтың бірінші сөзі: амин қышқылыньщ полипептидтік тізбекшесіне фенилала-ниннің енуін бақылайтын триплет урацил — урацил — урацил (УУУ) оқылды.
(7Л. Ниренберг, Г. Корана. және басқалардың зерттеулері ар-қасында қазіргі уақытта барлық амин қышқылдарына арналған триплеттер анықталған (5-таблица). Әрбір амин қышқылына бірнеше триплеттер — кодондар сәйкес келетініне, яғни кодтың өзгергеніне назар аударыңдар.
5-та блица
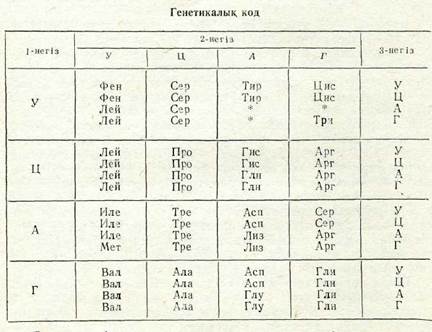
Е с к е р т у: Амин қышқылдарының аттары жалпы қабылданған қысқар-тулармен берілген.
* Амин қышқылдарының бірде-бірін кодпен анықтамайтын, полипептид-тік тізбек синтезінің аяқталғакын білдіретін триплеттер.
Белгілі бір амин қышқылын кодпен анықтайтын бірнеше триплетте, әдетте, екі негІз ортақ болады да, үшіншісінде айыр-машылық болады, бірақ көбінесе сол (пуринді немесе пирими-динді) топқа жатады. Сонымен қатар, УАА, УАГ және УГА триплеттері нонсендер болып табылатыны — олар амин қыш-қылдарының бірде-бірін кодпен анықтамайтыны, бірақ термина-торлар қызметін атқаратыны, яғни полипептидтік тізбек синтезі-нің аяқталуын анықтайтыны көрсетілген. АУГ кодоны и-РНК молекуласының бас жағында болған кезде, эукариоттарда поли-пептидтік тізбекке метиониннің, ал прокариоттарда — формил-метиониннің қосылуыи бақылайтын жағдайларда (бактериялар мен фагтарда сондай-ақ ГУГ триплеті болуы да мүмкін) белок синтезін жүргізуші механизм ролін атқарады.
Генетикалық кодты зерттеу жалпы биологиялық зор маңызы бар жаңалык ашуға мүмкіндік берді: кодтын, белгілі универсал-дылыгы анықталды. Бірдей триплеттер әр түрлі организмдерден (бактериялар, сүт қоректілер клеткасы) алынған клеткасыз сис-темалардағы бірдей амин қышқылдарын кодпен анықтайтыны көрсетілген. Код вирустар, балдырлар және теңіз кірпісі үшін универсалды болып шықты. Код, сірә, ежелгі әрі консервативті филогенездік механизм болса кереку'
Генетикалық кодтың ашылуы және белоктың іп үііго синтезі ғылымдағы тірі табиғаттың бірлігі жөніндегі мәселені мүлде жаңаша қойды. Бүл жаңалық белокты қолдан синтездеу әдісін меңгеру және бүл арқылы тірі материяның мәиіи неғұрлым те-рең танып білу жолындағы маңызды қадам болып табылады.
ЦЗонымен, ДНК — клеткадағы түкым қуалау информациясын тасушы. Онтогенезде информацияның жүзеге асуы үнемі жүріп отырады, бірак и-РНК синтезі митоздық циклдегі белгілі бір кезеңге, яғни хромосомалар деспиральданған күйде болатын ин-терфазаға сәйкестендірілгещ Бүл процесті ооциттердің «лампа щеткалары» түріндегі хромосомаларынан деспиральданған учас-келер — пуфтер пайда болған кезде қос қанаттылардың алып хромосомаларынан бақылау ыңғайлы. Түқым қуалау информа-циясын бір үрпақтан екінші үрпақка тасымалдауды клеткалар-дын, бөлінуі процесінде ДНК жүзеге асырады. Бүл жағдайда хромосомалар тығыз спирализация күйінде болады (2-тарауды караңыз).
Алайда, ДНҚ бәрін де атқара а*лады деп есептеу дүрыс бол-мас еді. Ол ферменттер, нуклеотидтер және оған негіз қалайтын ДНК. міндетті түрде болатын жағдайларда ғана репликациялана (екі еселене) алады, өйткені оған көбеюдің матрицалы механиз-мі тән. Белоктар (ферменттер) мен бастапқы өнімдер болған кезде де ДНК қызмет атқара (и-РНК синтезі) алады.
3. ӨЗГЕРПШТІКТІҢ МОЛЕКУЛАЛЫҚ НЕГІЗДЕРІ
Мутацияның молекулалық механизмі. Егер генетикалық ерекшелік ДНК молекуласындағы иуклеотидтер жұптарының жүйелілігімен анықталатын болса, мутацияның пайда болуы нуклеотидтер жүйелілігінің бүзылуымен байланысты болуы тиіс.
Мүндай өзгерістердің төрт түрі бар деп жорамалдауға бола-ды: нуклеотидтердің біреуінің немесе көбінің алмасуы, нуклео-тидтер жүптары жүйелілігінің өзгеруі, нуклеотидтердің бір не-месе көптеген жүптарының екі еселенуі, нуклеотидтердің бір немесе көптеген жүптарының түсіп қалуы.Клеткада белгілі бір химиялық заттардың кездесуі нуклеин
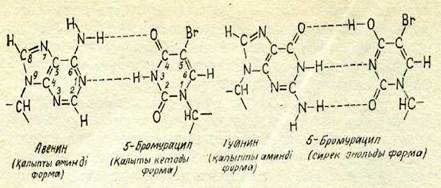 |
82-сурет. Аденин мен гуаниннің 5-бромурацилмен жұптасу схема-сы.
Қышқылдарына алғы шарт болатын заттардың қалыпты түзілуін тежеуі мүмкін. Мысалы, пуриндер синтезін тежейтін азосерин күшті мутаген болып саналады: уретан да пиримидиндер синте-зіи тежейтін күшті мутаген болып табылады.
Мутагендік әсер ДНҚ-да негіздер аналогтарын қосып алу процесі жүруіне келіп соғуы мүмкін. Бактериялар, фагтар мен адам тканьдері культурасының ДНК-дағы аденинге тиминнің орнына 5-бромурацил және азотты негіздердің басқа да туынды-лары қосылуы мүмкін екендігі дәлелденген. 5-бромурацил кейде, бірақ сирек жағдайда гуанинмен жүптасады. Жүптасудың екі схемасы да 82-суретте көрсетілген. Егер қателесіп, тиминнің орнына аденинге 5-бромурацил қосылатын болса, онда ДНК-ның келесі репликацияларында 5-бромурацил кейіннен цитозинмен қалыпты түрде жүптасатын гуанинді қосып ала алады. Осының нәтижесінде А — Т негіздерінін, жүбы Г — Ц жұбына алмасуы мүмкін. Басқа жағдайда Г — Ц жүбын А — Т жұбы осылайша алмастыруы мүмкіы. Алмасу мүкіндіктерінщ бүл схемасы 83-суретте келтірілген. Күшті мутаген—азотты қышқыл негіздер-ді аминсіздендіру жолымен әсер етеді. ДНК-де гуанин, одан соң цитозин оңай аминсіздендіріледі, ал аденин бәрінен де қиын аминсіздендіріледі. Негіздердің алмасуы А — Т+±Г — Ц осының нәтижесі болып табылады.
Сірә, сыртқы ортаның нуклеотидтер жүйелілігінің «қателігі-не» әкеп соғатыи кез келген факторлары мутагендер болуы тиіс болар.
Акридии — иприттердің бактерияларға әсерін зерттеу нәти-жесінде мутагендер әсері ерекшелігінің дәлелдемелері алынды.
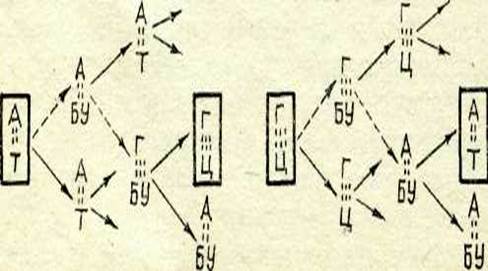 |
83-сурет. Негіздер жұбының алмасуы:
А— Г-ның Г— Д-ге жәие Г—Ц-ның А — Т-га ауысуы. Пунктир стрелка қате жұптасуды көрсете-ді. Нуклеотидтер арасындағы нүктелі линияілар саны су-тектің байланыстар санына сай келеді.Бұл мутаген негізінен «ілгерілемелі оқу» мутацияларын индукциялайтын болып шықты, яғни жекелеген нуклеотидтің жойы-луына немесе қосылуына себепші болады. Басқа да мысалдар белгілі. Алайда бұл мәселе әлі де шешілмеген күйде калып ке-леді.
Ультракүлгін сәулелердің ДНК ерітінділері мен бактерия-ларға әсерін зерттеуде, олар ДНК тізбекшесінде қатар орналас-са, тиминнің екі негізі арасында байланыс түзілуіне себепші бо~ латыны анықталды. Мүндай байланысқан негіздер тимин димері деп аталды. Димерлер неғұрлым көп түзілсе, бактериялар со-ғұрлым көп қырылғаы. Егер сәулелендіруден кейін бактерия-ларды жарықта қалдырса, олар аз қырылатын болған. Байқа-сақ, клеткада зақымданудың зардабын жоя алаіын механизмдер бар екен. Клеткадағы зақымдануды қалпына келтіру репарация деп аталды. Репарацияның дорепликативті репликация актісі кезінде және одан кейін өтетін (пострепликативті) репарация деген топтарын ажыратады. Дорепликативтік репара-ция механизмдерінің бірі — фотореактивация — клеткада жа-. рықта фотореакциялаушы фермент тиминдер арасындағы байланыстарды үзуге қабілетті болады. Р. Сетлоудық осы қүбылыс-ты зерттеген лабораториясында сәулелендіру нәтижесінде тиминдер димерлері ғана емес, сондай-ақ басқа да димерлер түзіле алатындығы көрсетілді. Репарация ферменттері өсімдіктерден, жануарлар мен адамдардан табылды. Бұл қарастырылып отыр.ған қүбылыстың универсалдылығын бідіреді. Дорепликативтік' репарацияның тағы бір түрі эксцизиялыц репарация деп аталды. Ол ультракүлгін сәулелердің әсерінен ғана емес, сондай-ақ қа-рақғыда иондаушы радиациялар мен химиялық мутагендердің-әсерінен болатын зақымдануларды да емдеп жазады. Қараңғы-лық репарацияның механизмі көп сатылы болады. Ол түзілген димерді «ойып алудан» басталады және эндонуклеаза ферменті-нің қатысуымен жүреді. Осыдан кейін ДНК-ның зақымданған жіпшесінің оны қоршаған негіздері (1 димерге бірнеше ондықтан мыңға дейін) ойып алынады. Процесс экзонуклеаза ферментінің жәрдемімен жүреді. Бұдан кейін ДНК-полимеразаның жәрдемімен ДНК-ның репарациялық синтезі жүреді. ДНК молекуласының зақымданбаған жіпшесі матрица қызметін атқарады. Зақымданған учаске мен ДНК-ның жаңадан синтезделген тізбе-гінің үшы лигаза ферментінің жәрдемімен біріктіріледі.
Пострепликативтік репарация димерлерді «ойып алумен» байланысты емес және ол репликация актісі кезінде жүзеге асады. ДНК репликациясы тізбек бойымен димерге дейін жәнеодан кейін де жүре береді, нәтижесінде димердің қарама-қарсысында ДНК-ның тізбегінде зақымдану пайда болатыны соңғы кезде анықталды. Бүдан соң сіңлілі дублекстер арасындағы рекомби-нация процесінде түзілген зақымдану зардабын жою, яғни за-қымдану репарациясы жүзеге асады. Адам клеткаларындағы репарацияның маңызы туралы, мысалы, репарация системасы бүзылған адамдарда пайда болатын зілді ауруларға қарап айтуға болады: пигменттік ксеродермада күн сәулесі әсер еткенде тері зақымданады, ол әдетте тері рагына ұшырап өліммен аяқта-лады.
Мәселен, эволюция процесінде келесі ұрпақта тұқым қуалау информацияоының константтылығын қамтамасыз ететін меха-низмдер пайда болды, Алайда эволюция процесінде особьтардың алуан түрлілігін, олардың өзгергіштігін қамтамасыз ететін ме-ханизмдер де түзілді. ДНК молекуласындағы нуклеотидтер жүйелілігінін, өзгеруіне, яғни мутацияға әкеп соғатын репликация қателІктерін; и = РНК-нық жаңа молекулаларының пайда болу-ына және осының салдары ретінде организмде жаңа белок мо-лекулаларының пайда болуына әкеп соғатын транскрипция қат-ліктерін; жаңа белоктардың пайда болуына әкеп соғатын трансляция қателіктерін осындай механизмдер қатарына жатқызуға болады. Репарация механизмі де қателік жіберуден құр емес, сондықтан мутация көзі болуы мүмкін. Мутациялық процестің физиологиялық гипотезасын М. Е. Лобашев 1947 жылы-ақ тұ-жырымдаған болатын, бұл гипотезаға сәйкес мутация дегеніміз клеткада пайда болған зақымданудың теңсіздік репарациясы-ның салдары болып табылады. Мутациялық процеске берілген бүл көзқарас қазіргі уақытта эксперименттік жолмен дәлелде ніп отыр.
Кроссинговердің молекулалық механизмі. Тұқым қуалау ин-формациясын жеткізуші нуклеин қышқылдары жөніндегі түсінік тіркескен гендер рекомбинациясының механизмін, яғни кроссинговер механизмін түсіндіретін бірқатар гипотезалар ұсынуға мүмкіндік берді. Бүл гипотезалардың әр түрлі нүсқалары 1963 жылы К. Уайтхауз тұжырымдаған идеяға келіп соғады.
Әрбір хроматидте ДНҚ-ның бір молекуласы берілген деп есептегенде екі гомологты хромосомалар учаскелері ал-масуының бүкіл процесін бірнеше кезеңге бөлуге болады: ДНҚ-ның екі молекуласының жақын, бірақ бір-біріне ұқсас емес нүк-телерінде бір тізбектін, үзілуі (84-сурет, 2); ДНК-ның үзілу учаскесіне жақын жердегі комплементарлы тізбекшелер арасындағы сутектік байланыстардың үзілу және ДНК мөлекуласының жаңа биспиральды «будандық» құрылымының түзілуі (84-сурет, 3); айрылып кеткен тізбекшелер орнында ДИҚ-ның жергілікті синтезделуі (84-сурет, 4); жаңадан синтезделген тіз-бекшелердің жан-жаққа қашықтауы (84-сурет, 5) және олардың комплементарлы жалғасуы (84-сурет, 6); рекомбинацияға үшы-рамаған тізбекшелердің үзілуі (84-сурет, 7); және үзілген үшта-рының түйықталуы (84-сурет, 8).
Процестін, барлық кезеңдері мейоздың ерте профазасында, яғни хромосомалардың редупликациясы аяқталған соң ғана өте-ді. Әр түрлі объектілерден алынған эксперименттік фактілердің кепшілігі үсынылып отырған рекомбинация механизмін қанағат-танарлық дәрежеде түсіндіреді.
Уайтхауз үсынған модель бірден-бір модель емес. Р. Холлидей үсынған кроссинговер механизмінің моделі де қолда бар ге- 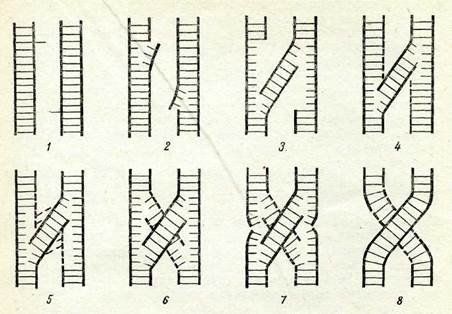
84-сурет. Рекомбинацияның молекулалық деңгейде мүмкін болатын механизмі (бастапқы үзілу нүктелері стрелкалармен көрсетілген).
Тұтас сызыңтар — ДНК спиральдарының жалац ңабат тізбекшелері: пунктир сызьщ-тар — ДНК-ның жаңадан синтезделген тізбекшелері: 1 — 8 — рекомбинация кезеңдері.нетикалық фактілермен әжептәуір үйлеседі. Алайда бұл мәселе-ні түпкілікті шешілген деп есептеуге болмайды. Рекомбинация процесі репликация (мейоз профазасындағы ДНҚ синтезі, 3-та-рауды қараңыз) және репарация процестерімен тығыз байла-нысты екенін атап көрсету керек. ДНК молекуласы жіпшелерінің үзілуі және кроссинговер кезінде үзілген ұштарының бірігуі ре-парациялық ферменттердің көмегімен іске асады.
4. ГЕНЕТИКАЛЫҚ ИНЖЕНЕРИЯ
Трансформация, трансдукция және кері транскрипция, бакте-риялардан плазмидтер мен эписомаларды табу, генетикалық кодты, белок синтезінің механизмін талдау секілді құбылыстар-ды ашу клеткалардың молекулалық реконструкциясы сияқты міндеттерді алға қоюды шындыққа айналдырды, ал бұл саладағы іс жүзіндегі табыстар генетикада жаңа бағыт — генетикалық инженерияның тууына себепші болдьиГендік инженерияның бо-лашағы шексіз: адамдарды «ауру» гендерді сау гендермен ал-мастыру жолымен емдеу, ауа азотын сіңіретін өсімдіктерді шы-ғару, белгілі бір өнімділік беретін жануарларды шығару және т. с. с. Қазіргі генетиканың табыстары өте зор. Г. X. Қорана ала-нинді транспорттық РНК-нын, клеткада қызмет атқармаған генінің және ішек таяқшасының клеткасына енгізгенде калыптьг қызмет атқарған тирозинді транспорттық РНК генінің «жартылай химиялық» синтезін жүзеге асырды. т-РНКгпир геніндегі. нуклеотидтердің жүйелілігі 1967 жылы?чықталған болатын. Генкің синтезі бірнеше кезеңде жүзеге асырылады. Алдымен нуклеотидтердің шағын жүйелілігі (10—15) сиытезделді. 40 шақ-ты фрагмент алынған соң оларды ферменттердің (лигаза) жәр-демімен біріктіріп, 126 нуклеотиді бар молекула — геннің өзІ және екі реттеуші учаске — геннің жүмысын инициалаушы про-мотор (52 жүп) және ген жүмысының аяқталуын анықтайтын терминатор (нуклеотидтердің 21 жүбы) алынды.
Кері транскрипцияның ашылуы генетиктерге генді синтездеу-дің жаңа әдісін берді, мүның жәрдемімен адам, кролик, тышқан мем үйрек гемоглобинінің ғеидсрі, сондай-ақ, басқа да г








