 2015-06-28
2015-06-28 5566
5566Әдетте әр түрлі жыныстың особьтарын бір-бірінен айыратын белгілер бастапщы және қосымша жыныстыщ белгілерге бөліне-ді. Организмнің гаметалардың түзілуін және үрықтану проце-сінде олардың қосылуын қамтамасыз ететін морфологиялық жә-не физиологиялық ерекшеліктерін бастапқы белгілерге жатқы-зады. Олардың қатарына, мысалы, жоғары сатыдағы жануар-лардың гонадалары (жыныс бездерІ), жыныс жолдары мен сыртқы гениталийлері (жыныс мүшелері), жоғары сатыдағы өсімдіктердің андроцейлері (аналықтарының жиынтығы) мен гинецейлері (аталықтары) жатады. Организмнің гаметогенез, шағылысу және үрықтану процестерін тікелей қамтамасыз ет-п£Йтін, бірақ жыныстық жолмен көбеюде белгілі бір қосымша роль атқаратын белгілері мен қасиеттёрін қосымша белгілерге жатқызады. Оларға балықтардын жүзу қанаттарының, қүстар-дың қауырсындарының, сүт қоректілердің көкірек бездерінің, жоғары сатыдағы есімдіктердің буын аралығының үзындығы мен гүлдеу мерзімдерінің ерекшеліктері және т. б. жатады.
Кейбір төменгі сатыдағы және көптеген жоғары сатыдағы есімдіктер. мен жануарларда аталық және аналық жыныс особь-тарының айырмашылығы — жыныс диморфизмі кездеседі. Жо-ғары сатыдағы есімдіктердің 5 мыңға жуық түрі қос үйлі. Жо-ғары сатыдағы жануарларда қос жынысты (гермафродитті) формадар нормада кездеспейді. Барлық жағдайларда да айыр-машылық жануарлардың морфологиялық, биохимиялық, физио-логиялық белгілеріне, экологиясына, мінез-қылығына және т. б. қатысты болады. Алайда бір клеткалы организмдердің төменгі сатысында копуляцияға қатысатын особьтардың айырмашылы-ғы кезге көрінбеуі және физиологиялық ерекшеліктерге ғаиа қатысты болуы мумкін, сондықтаи олардың жынысын шартты түрде «+жыныс» немесе «—жыныс» деп белгілейді. Екі типтің арасындағы айырмашылық көбінесе сандық сипатта болады, сондықтан кептеген бір клеткалы балдырлардан (мысалы, хла-мидомонадалар), кейбір төменгі сатыдағы саңырауқұлақтардан немесе инфузориялардан ішінде клеткалар копуляциясы бол-майтын, ал линиялар арасында ол жүзеге асатын бірнеше ли-нияны бөліп көрсетуге болады. Көптеген линияларды «+» және «—» типтерге топтастыруға болады, бірақ тип ішінде де кейде клеткалар копуляциясы болатын әр түрлі линиялар бар екенді-гі анықталды.
Демек, мүнда салыстырмалы сексуалдылық туралы айтуға болады.
Жыныс диморфизмі неліктен пайда болды? Бүл сурауға жа-уап беру үшін жыныстық процестің биологиялық мәні тұқым қу-алайтын өзгергіштіктің жалпы қорының артуына әкеп соғатын түқым қуалау информациясының қайта комбинациялануында екенін еске түсіру керек. Бүған алғаш рет Ч. Дарвин назар ау-дарған. Ол шағылыстырудың биологиялық пайдалылығы заңын түжырымдап, ол арқылы екі жыныстың шығуын түсіндірді.
2. ЖЫНЫСТЫ АНЫҚТАУДЫҢ ХРОМОСОМАЛЫҚ ТЕОРИЯСЫ
Дара жынысты организмдерде жыиыстардың арақатынасы 1: 1-ге ж.ақын, яғни еркектері мен үрғашылары бірдей қатынас-та кездесетінін адамдар өте ертеден-ақ байқағаи.
Мүндай 1: 1 ажырау талдаушы шағылыстыруға Аа~Хаа тән екеніне кезінде Мендель назар аударған болатын. Жыныстардың біреуі гомозиготалы, екіншісі — гетерозиготалы болуы тиіс де-ген жорамал айтылды. Бүл гипотезаның дүрыстығын көрсететін алғашкы зксперименттік дәлелді К. Корренс тапты. Вгуопіа туы-сының ішінде қос үйлі (В. сііоіса) және бір уйлі (В. аіЬа) турлері ба&. Кос үйлі түрдің аталық жәңе аііалық өсімдіктерінің жыныс-тары қалай тұқым қуалайтыиыи анықтау үшін оларды бір үйлі түрлермен шағылыстыру жүргізді. Аналық өсімдіктер үрпағын-да тек аналық, ал аталық өсімдіктер үрпағында — жартысы аналық және жартысы аталық есімдіктер болып шықты. Осы-дан келіп, Вгуопіа-иъщ аналық өсімдіктері гомозиготалы, ал ата-*лық есімдіктері гетерозиготалы.. 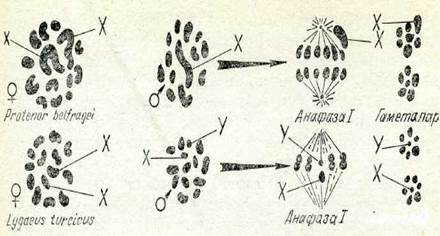
Г&штал&р
85-сурет. Аталықтар мен аналықтар кариотиптері және гетерогаметалық жыныс гакеталарыныц хромосома жиынтықтары..
Жынысты анықтайтын бірдей гаметалар түзетін жынысты гожогаметалы, ал әр түрлі гаметалар түзетін жынысты — гете-рогаметалы жыныс деп атайды.
Жоғарыда айтылғанындай (8-тарауды қараңыз), мүндай қорытындының дұрыстығына шешуші дәлелді цитологтар тап-ты. Өткен ғасырдың соңында Ьу§;аеиз қандаласынан спермато-генезді зерттегенде Х-хромосомалы және У-хромосомалы екі сортты гаплоидты сперматоциттер II сипатталды (85-сурет). Ұрғашыларының жүмыртқа клеткаларында еркектерімен бірдей 6 аутосомалардан басқа, міндетті турде Х-хромосома болған. Ргоіепог деп аталатын басқа қандалада да аталық жыныс ге-терогаметалы болып шықты. Бірақ бұл түрде 6 аутосомалар-дан басқа, сперматоциттердің жартысында Х-хромосома бол-ған, ал жартысында ол болмаған (85-суретті қараңыз).
X- және Ғ-хромосомалардың жынысты анықтауға қатысы бар деген жорамал айтылды, оларды жыныс хромосомалары деп атады. Т. Морган мен оның қызметкерлері жыныспен тір-келғен белгілердің түқым қуалаушылығын зерттеуде бүған эксперименттік дәлелдер тапты (8-тарауды қараңыз). Жыныс-ты аныщтаудыц хромосомалар теориясы алғаш рет осылайша тұжырымдалды.
Бұл жаңалық онан әрі цитологиялық зерттеулерді күшейту-ге жол ашты. Жыныс хромосомалары көптеген организмдерден табылды. Өсімдіктердін, арасынан жыныс хромосомалары ал-ғаш рет бауыр мүгінен Зрһаегосагриз сипатталды. Олар ме-ландриум, қымыздық, элодея, қүлмақ және т. с. с. жоғары са-тыдағы әсімдіктерден де белгілі. Жануарлардан олар көптеген насекомдар, қүстар, сүт қоректілер үшін сипатталған. Адамның да жыныс хромосомалары сипатталған.
Л^ыныс хромосомаларын зерттеу олардың аутосомалардан генетикалық ғана емес (8-тарауды караңыз), сондай-ақ цитоло-гиялық түрғыдан да айырмашылығы барын көрсетті. Жыныс хромосомалары гетерохроматинге бай келеді (2-тарауды қара-ңыз). Олардың репликациясы аутосомалармен асинхронды өте-ді, ал гомогаметалы жыныста Х-хромосомалардың бірі қалғандарынан кешірек репродукцияланады. Мейозда олар көбінесе күшті спиральданады (гетеропикноз). Гетерогаметалы жыныс-тың жыныс хромосомалары (гетероморфты жұптар) коныога-цияланбайды немесе ішінара ғана коньюгацияланады, бұл оның жекелеген учаскелерінің ғана гомологты болатынын көрсетеді.
8-тарауда айтылғанындай, X- және К-хромосомалар ажыра-ғанда редукциялық бөліну кезінде бірі Х-хромосомалы, екінші-сІ У-хромосомалы әр түрлі 2 клетка түзіледі, демек гетерогаме-талы жыныс түзетін X- және У-хромосомалы гаметалардың аракатынасы дәл 1: 1 болады. Егер клеткада бір Х-хромосома болса, гаметалардың екі сорты дәл осылайша түзіледі: бүл жағ-дайда гаметалардың 50%-нің Х-хромосомасы болады, ал 50%-інің /Ү-хромосомасы болмайды. Гомогаметалы жыныс түзе-тін гаметалардың бәрі бірдей, әрі оларда Х-хромосома болады («гомогаметалы» деген атаудың өзі де осыны көрсетеді). Ұрық-танудың нәтижесінде еркектер мен ұрғашылар бірдей мөлшер-де пайда болады. Басқаша айтқанда, жынысты анықтаудың хромосомалық механизмі жыныстардың арақатынасын 1: I ша-масында сақтайтын өзін-өзі реттейтін керемет механизм болып табылады. Әр түрлі организмдердің жыныс хромосомаларына жасалған талдау жынысты хромосомалық анықтаудың әр түрлі типі бар екенін көрсетті. Олар ХОжәяе ХҮ тип деген атауларға ие болды. Аталық жыныс та, сондай-ақ аналық жыныс та гете-рогаметалы жыныс бола алады. Қазіргі кезде жыныс хромосо-маларыиың неғұрлым күрделі комплекстері де сипатталған, бі-рақ олардың біз атап өткен жыныс хромосомаларынан принцип-тік айырмашылығы жоқ,
Фактілердің жиналу дәрежесіне қарай жынысты анықтау-дың хромосомалық теориясы дәлелдене түсумен қатар, кейбір қиыншылықтарға да кездесті. Жыныс хромосомалары жыныс индикаторлары, жыныстық қосымша белгілер емес пе екен де-ген сүрау ашық күйде қалды.
Дрозофиланын, Бриджес тәжірибелерінде жыныс хромосома-лары ажырамауының нәтижесі ретінде алынған айрықша особь-тарына жасалған талдау (8-тарауды қараңыз) аутосомалардан басқа АлХҒ-хромосомалары (ХХҮ+АА) бар особьтар үрғашы-лар, ал ХО~\-АА особьтары — еркектер болып табылатынын көрсетті. Бүл фактілер жыныс хромосомалары мүлде жыныс индикаторлары емес екендігіке сенімді дәлел болады. Алайда, ХҮ+АА және ХО + АА особьтары еркек, ал XX-\-АА және ХХҮ-\-АА особьтары үрғашы болса, олар қалайша жынысты анықтайды? Сірә, мұндағы мәселе алғашқыда көрінгеніндей болмаса керек.
3. ЖЫНЫСТЫ АИЫҚТАУДЫҢ БАЛАНСТЫҚ ТЕОРИЯСЫ
Бриджес гипотезасы. К- Бриджес дрозофиладан хромосома-ларвшың триплоидтық жиынтығы ЗХ+ЗЛ болып келген бірнеше үрғашыларын тапты. Оларды диплоидты еркектерімен ХҮ+2А шағылыстырғанда морфологиялық, цитологиялық және генети-калық зерттеулер жыныс хромосомалары мен аутосомалар жи-ынтықтарының арақатынасы түрліше болып келген особьтар-дың сегіз типі бар ұрпак алынды: ЗХ:ЗА; 2Х:2А; (2Х+Ү):2А; 2Х:ЗА; (2Х+Ү): ЗА; ХҮ: 2А; ЗХ: 2А; ХҮгЗА. Анеуплоидты шыбындардың пайда болуы триплоидты үрғашыларда қалыпты коньюгацияның бұзылуы мен мейоздық хромосомалардың ажы-рауына байланысты (13-тарауды қараңыз).
Аналық зиготада аутосомалар жиынтығынын, басым болуы (2Х: ЗА) жынысы аралықта дамитын особьтардың, яғни интер-секстердің түзілуіне әкеп тіреді. Х-хромосомалар санының кемуі (X: 2А) аталық жыныстың дамуына себепші болды. Бірақ бір /Ү-хромосомаға келетін аутосомалар жиынтығының саны үшке дейін артатын болса (Х:ЗА), онда асқын еркек еркектік белгілері пштертрофияға ұшыраған организм дамиды. Ол стеркльді бо-лады. Қерісінше, аутосомалардың диплоидты санына шаққанда Х-хромосомалар сакының артуы (ЗХ: 2А) жұмыртқа безі қа-лыпты дамымаған және жыныстық басқа да бұзылған белгіле-рі бар асқын үрғашы особьтың дамуына жеткізеді.
Осы тәжірибелердің негізінде Қ. Бриджес дрозофилада ана-лық жынысты екі Х-хромосоманың және аталық жынысты — ХҮ хромосоманың болуы емес, жыныс хромосомалары саны мен аутосомалар жиынтығының аракатынасы анықтайды деген қо-рытындыға келді. Аналық тенденцияның гендері негізінен Х-хромосомаларда, аталық тенденцияның гендері — аутосома-ларда шоғырланған. Бүл хромосомаларының балансы (немесе жыныстық индексі) X: А—\ болатын барлық особьтар аналық-тар болып табылатындығынан, ал X: 2Л = 0,5 қатынасы ата-лықтарды беретіндігінен көрінеді: қатынасы 1-ден 0,5-ке дейінгі хромосомалар балансы интерсексуальдылықты анықтайды. Үш Х-хромосоманың аутосомалардың екі жиынтығына катынасы ЗХ:2А=\,5 асқын үрғашының дамуына себепші болады. Кері-сінше, бір Х-хромосомаға аутосомалар жиынтығы мөлшерінің артуы уҮ."574 = 0,33 асқын еркектің дамуын анықтайды. Бұл формаларды суперсекстер деп атайды. Дрозофиладағы У-хро-мосома жынысты анықтамайды.
Дрозофилада жыныс көптеген гендердің балансымеи анық-талатындығына, ал Х-хромосома аналық жынысты, аутосома-лар аталық жынысты анықтайтынына дәлелдер табу үшін қо-сымша үйлесімді тәжірибелер жүрғізілді. Хромосомалық қайта қүрылу (дупликациялар) әдісімен екі А'-хромосомамен ауто-сомалардың үш жиынтығынан басқа Аг-хромосоманың үзынды-ғы әр түрлі қосымша учаскелері бар особьтар алына бастады. Мұндай учаскелердің ұзаруына қарай интерсекстер күтілгенін-дей, барған сайын үрғашыларға үқсай бастады. Сонымен қатар, триплоидты үрғашылардың бірқатар үрпағында интерсексуал-дылықты кәрсетуге, яғни аналык немесе аталық типтің иитер-секстерін беру қабілетін анықтауға бағытталған селекция жүр-гізілді. Мүнда бүл екі линияның триплоидты үрғашыларындағы хромосомалардың арақатынасы интерсекстердегі хромосомалар-дың арақатынасындай бұрынғы қалпында сақталды -■- ЗХ: ЗА; ал ұрпақтарда интерсексуалдылықтьщ көрінісі селекцияның ба-ғытына сәйкес өзгерді.
Жынысты анықтаудағы баланстық теориясының универсал дығы. Генетиканың соңғы жылдары жиыақтаған көптеген та-быстары жынысты анықтаудағы баланстық теориясын тұжы-рымдауға мүмкіндік берді, бұл теория бойынша особьтың жы-нысы геномның кез келген хромосомасында орналасқан, аталық және аналық жынысты детерминдеуші гендер балансымен анықталады. Гендер балансын қатаң сақтайтын және жыныста-ры жөнінен айқын жіктелген особьтардың пайда болуын қамта-масыз ететін механизм—жыныс хромосомаларынын, әсер ету ме-ханизмі болып табылады. Мәселен, қазіргі уақытта адамның кемінде 6 гені (Х-хромосомадан 3 және У-хромосомадан 3 ген) сипатталған, олардың өзара әсер етуі, балансы особьтың жыны-сын анықтайды. Алайда, дрозофиладан өзгеше, адамның У-хро-мосомасы (Н —У антигені) аталық жынысты анықтауда шешу-ші роль атқарады. У-хромосома болмаған және Х-хромосоманын, кез келген саны болғанда особь фенотип бойынша ұрғашы ре-тінде анықталады. Тұт көбелегінде У-хромосома Х-хромосома-ның кариотиптегі санымен еселену дәрежесіне қарамастан, ана-лық жынысты анықтай отырып балансты өзгертеді.
Жынысты анықтаудағы баланс теориясын жоғары сатыдағы ©сімдіктерге де қолдануға болады. Дрема (Меіапйгіит) есімді-гінде жыныс ең алдымеи X- және У-хромосомалардың балансы-мен анықталады. Аутосомалар санының өзгеруі ешқандай нәти-же бермейді. Бірақ тіпті У-хромосомасынан айрылған аналық өсімдіктердің де аталық потенциясы болады. Мәселен, аналық есімдіктер қара күйемен зақымданғанда гүлде ■ аталық өсіп-жетіледі, яғни гүл морфологиялық жағынан гермафродитті бо-лады. Особьтардың жынысын жыныс хромосомалары емес, бірнеше гендердің өзара әсер етуі анықтайтын жағдайлар да мәлім. Мысалы, шаншарларда (НаһгоЬгасоп), бал араларының оған жақын түріндегі сияқты үрықтанған жүмыртқадан үрға-шылары, ал үрықтанбаған жүмыртқадан еркектері дамиды. БІ-рақ шаншарларда туыстық жағынан жақын түрлерді шағылыс-тырғанда үрықтанған жүмыртқадан ерксктері дамитын жағдай-лар да сипатталған. Ұрықтану процесінің болғанына немесе бол-мағанына қарамастан, шаншардыц аналық жынысын екі фак- а- тордың гетерозиготалық =^> күиі, ал аталық жынысын — ХаХ~\~ Х^гХ^Ха %'~т~ ү-[-уһ
гомо- ^т^-, х+х~ь немесе гемизиготалық күйі анықтайтынын генетикалық талдау көрсетті. Қазіргі кезде бал араларында да үрықтанған жүмыртқадан кейде еркек ара-лар шығуы мүмкіндігін көрсететін мәліметтер алынды. Мүмкін оларда да жынысты анықтаудың осыған үқсас механизмі бар болар.
Жыныс хромосомалары жоқ және жынысы гендер арқылы аныкталатын басқа да өсімдіктер мен жануарлар сипатталды.
Жынысты аыықтаудағы баланс теориясы қазіргі кезде көп-шілік қабылдаған теория болып табылады. Ол барлық дара жынысты организмдер мен олардың гаметаларының генетика-лық қамтамасыз етілген потенциалдық бисексуалдылығын көр-сетеді. Онтогенездегі гендер балансын сақтайтын механизмдер түрліше болуы мүмкін.
Жыныс хроматині. М. Барр мен Ч. Бертрам 1949 жылы мы-сықтың нейронын зерттеуде клетканың интерфазалық ядросын-да кейбір бояғыш заттармен жедел боялатын түйіршіктер бо-латынына, оның үстіне ол ұрғашы мысық клеткасының ядро-сында бар да, еркектерінде жоқ болатынына назар аударды. Мүндай түйіршіктер көптеген жануалардан және оның гетеро-гаметалылығына қарамастан әрқашан да аналық жыныста кез-десетін болып шықты. Клетка ядросындағы бүл қүрылым жы-ныс хроматині немесе «Барр түйіршігі» деп аталды (VI табл, Б). Сүт қоректілердің үрғашыларының интерфазалық ядроларын-да жыныс хроматині болуын Х-хромосомалардың бірінің гете-ропикноздық күйімен байланыстырады, хромосоманың бүл инактивациясы, сірә, үрғашы және еркек особьтар клеткала-рындағы жыныс хромосомалары мен аутосомоларынын, балан-сын теңестіруші механизм болса керек (29-тарауды қараңыз). Еркектер мен ұрғашылардағы гендер балансын теңестірудің басқа да механизмдері белгілі, мысалы, дрозофиладағы еркек-терінің Х-хромосомасындағы гендердің гиперактивтілігі.
4. ОРТА ЖАҒДАЙЛАРЫНЫҢ ЖЫНЫСТЫ АНЫҚТАУДАҒЫ РӨЛІ
Орта жағдайларының жынысты анықтаудағы ролі туралы мәселені арнайы қарастыру керек. Белгілі дара жынысты орга-низмдердің кепшілігінде орта жағдайлары особьтың жынысын бақыламайды. Жыныс тек генетикалық механизм арқылы анық-талады.
Аздаған теңіз омыртқасыздарында, мысалы, Вопеіііа ьігійіз қүртында особьтың жынысын сыртқы орта анықтайды. Қүрт-тың үрғашылары ірі, микроскоптық мөлшері ғана бар еркекте-рі үрғашыларының жатырында паразиттік тіршілік етеді. Жы-ныстық түрғыдан алғанда индифференттік личинкалар үрғашы-ларының тұмсығына бекінуі мүмкін, бұл жағдайда олар еркек особьтарға айналады, ал еркіндікте тіршілік етіп, үрғашы особьтарға айналуы да мүмкін. Егер үрғашысының тұмсығына қоныстанған личинканы оқшаулап, жеке өсірсе, ол интерсекс болып шығады. Аталық және аналық жынысты анықтайтын гендер тепе-теңдік күйінде болып және орта жағдайлары ба-лансты екі жыныстың біреуінің басым болуы жағына қарай ез-гертетін кезде жынысты бүлайша анықтау әдісі, сірә, үрпақ қалдырудың ең қолайлы мүмкіндігін қамтамасыз ете отырып, осы түр үшін барынша тиімді болса керек. Эволюциялық тұрғыдан алғанда дара жынысты жануарларда бәлкім, бұл өте қа-рапайым әдіс болар.
Жыныс белгілерінің оюыныс диффвренциациясы деп атала-тын онтогенезде қалыптасу процесін жынысты анықтаудан айы-ра білу керек. Корыта келе, тірі табиғат құрылымының барлық деңгейінде организмдер генетикалық жағынан бисексуальды болып табы-лады, яғни олардың екі даму мүмкіндігі болады, және жынысты анықтау олардың сақтау механизмі әр түрлі бола алатын гендер балансының нәтижесі деп айтуға болады. Жыныс хромосомала-рының өзін-өзі реттеу системасы неғұрлым кең таралған.
19-тарау ЖЫНЫС ДИФФЕРЕНЦИАЦИЯСЫ ЖӘНЕ ЖЫНЫСТЫ ҚАЙТА АНЫҚТАУ
Организмнің кез келген басқа белгілері секілді, жыныс бел-гілерінің дамуы да генотип және сыртқы орта факторлары ар-қылы анықталады.
Организмдер генетикалық бисексуальды болғандықтан. жы-ныстың дифференциациялануы процесі күрделі болады. Орга-низмнің бисексуальдық негізі сайып келгенде оның даму бағы-тын өзгертуге, яғни онтогенезде жынысты қайта анықтауға мүмкіндік береді.
1. ЖЫНЫСТЫҢ ДИФФЕРЕНЦИАЦИЯЛАНУЫ
Жыныс анықталған соң іле-шала дифференциациялану, яғ-ни жыныс айырмашылықтарының даму процесі: көбею система-ларының, сондай-ақ шағылысуды қамтамасыз ететін физиоло-гиялық және биохимиялық механизмдердің қалыптасу процесі жүреді.
Жануарлар эмбриондарындағы жыныстық түрғыдан алған-дағы өсе бастаған индифферентті гонадалардың қосарлы сипаты болады. Олар дифференциация процесінде аналық жыныс клеткалары дамитын сыртқы қабат — кортекстен (согіех) және аталық гаметалар дамитын ішкі қабат — медулладан (тесіиі-1а) түрады.
Жыныс дифференциация барысында гонада қабаттарының бірі дамиды, екіншісі басылады. Аталық жыныста кортекстік қабаттың қызметін тежейтін медуллалық ткань тезірек жетіле-ді, осының нәтижесінде гонадалар аталық безге айналады. Ана-лық жыныста кортекстік кабаттың жетілуі тездейді, осының нәтижесінде медуллалық қабаттың қалыптасуы тежеледі де, гонадалар аналық безге айналады. Осындай өзгерістерге сәйкес жыныс жолдары да дифференциацияланады, олар екі жыныс-тың особьтарында да бірдей болып салынады.
Көптеген жануарларда жыныстың дифференциациялану процесін гормондар қамтамасыз етеді, гормондарды эндокриндік бездер ғана емес, сондай-ақ жыныстық бастаманың кортекстік және медуллалық қабаттары, ал кейініректе — жыныс бездері де бөліп шығарады.
Жыныстың онан әрі дифференциациялануы, әсіресе қосымша жыныстық белгілердің дамуы да әр түрлі гормондардың әсері-мен өтеді. Гормондық секреция дәрежесін гендер, дәлірек айт-қанда, олардың балансы бақылап отырады. Жалпы баланста аталық жынысты анықтайтын гендердің басым болуы аталық гормондар активтілігінің артуына және аталық жыныстың диф-ференциациялануына, гендердің кері арақатынасы — аналық жыныстың дамуына әкеп соғады. Онтогенезде кейде бір жыныс-тың немесе екінші жыныстың гормондық секреция активтілігі-нің алмасуы интерсексуальдық формалардың дамуына әкеп ті-рейді.
Өсімдіктер гормондары — ауксиндер жоғары сатыдағы өсім-діктердің жыныс дифференциациясына айтарлықтай әсер етеді.
2. ОНТОГЕНЕЗДЕ ЖЫНЫСТЫ ҚАЙТА АНЫҚТАУ
Онтогенезде жыныстың табиғи немесе жасанды жағдайлар-да өзгеруі организмдердің тұқым қуалайтын бисексуальдылы-ғының ең жаксы дәлелі болып табылады.
Сүт қоректілерде түрлі жынысты егіздер дамуында кейде олардың біреуінің жынысы эмбриогенезде өзгереді. Мәселен, мүйізді ірі қараның түрлі жынысты егіздерінде еркек бұзау қалыпты ©сіп-жетіледі де, ұрғашы бұзау көбіне интерсекс бо-лып шығады. Мүндай жануарлар фримартиндер деп аталды; олар әдетте үрықсыз болады. Мүндай езгерістер аналық без-дерге қарағанда аталық бездердің қанға аталық гормондарды ерте бөліп шығара бастауына байланысты болады. Бүл жалғыз мысал емес. Особь онтогенезінде арнайы әсер етпей-ақ жыныс-ты қайта анықтаудың басқа да жағдайлары белгілі.
Алайда соңғы жылдары жынысты эксперименттік жолмен қайта анықтау мәселесі барған сайын зерттеушілер назарын аударуда. Онтогенезде жынысты толық қайта анықтаудың тама-ша мысалдарының бірі 1953 жылы Т. Имамотоның аквариум балықтарына жүргізген зерттеулері нәтижесінде алынды. Тәжі-рибе үшін қызыл реңді Я доминантты гені У-хромосомада, ал оның г рецессивті аллелі — Х-хромосомада болатын медактар (огугіаз Іатііез) алынды. Бұл жағдайда Хг Үя генотипі бар еркек балықтар әрқашан да қызыл реңді, ал ХгХя генотипі бар үрғашы балықтар — ақ реңді болады. Автор бүл белгінің тү-қым қуалауын бірнеше ұрпақтан тексерді. Мынадай $ Хг Хг X & Хг Үк шағылыстырудан әрдашан да ақ рең-ді ұр-ғашы және қызыл реңді еркек балықтар алынды.
Тәжірибеде жыныстық жағынан әлі де дифференциациялан-баған майда шабақтар 8 ай бойы азықпен бірге аналық жыныс гормонын (эстронды немесе стилбестрольды) қосымша алып
отырды. Осының нәтижесінде барлық балықтар (ақ және қы-зыл) фенотипі бойынша қалыпты аналық бездері және қосым-ша аналық жыныстық белгілері бар ұрғашы балықтар болып шықты. Олар қалыпты қызыл реңді еркек балықтармен шағы-лысуға қабілетті болды. Кызыл реңді аналықтар генотиптік аталықтар болғанын талдау көрсетті. Мұндай ұрғашыларды қалыпты еркектерімен шағылыстыру? Хг Үк X с? Хг Үя жынысы бойынша 1: 1 емес, 1 $ (Хг Хг):3 с? (2ХГ Үя және 1 Үя Үк) ажырауын берді.
Еркектердің Үя Ү® болуы шағылыстыру нәтижелерімен дә-лелденеді. Аталық гормон (метилтестостерон) әсер еткенде ак реңді балықтар, яғни ұрғашылар генотипі бар балықтар —• Хг Хг еркек балықтар болып шығады. Оларды қалыпты ұр-ғашылармен (Хг Хг) шағылыстырғанда ұрпақтарында тек қана ұрғашы балықтар алынады.
Бұл жалғыз мысал емес. Тритоннан (РіеигосІеіез хюаШіі), кейбір бақалардан (Хепориз Іаеуіз), көптеген балықтар мен бірқатар басқа да жануарлардан осындай нәтижелер алынды. Мұндай мәліметтер өсімдіктерден де алынды.Сонымен, организмдердің генетикалық бисексуальдылығы онтогенезде жыиыс дифференциациясының өзгеруіне, яғни особьтың жынысын қайта анықтауға мүмкіндік береді.
20-тарау. ЖЫНЫСТАРДЫҢ АРАҚАТЫНАСЫ ЖӘНЕ ОНЫ РЕТТЕУ ПРОБЛЕМАСЫ
Жоғарыда айтылғанындай, 1: 1 арақатынасын қамтамасыз ететін, жынысты анықтаудың генетикалық механизмі болады.
Жыныстардың генетикалық анықталған арақатынасын ал-ғашқы арацатынас деп атайды. Алайда даму процесінде аталық және аналық зиготалардың тіршілік қабілеттілігі әркелкі бо-луы, жынысты қайта анықтау және басқа себептердің салда-рынан жыныстардың арақатынасы өзгеруі мүмкін. Жеке даму процесінде әр түрлі факторлардың әсерінен болған арақатынас өзгерісін жыныстың цосымща аращатынасы деп атайды. Жы-ныстың қосымша арақатынасы көбінесе аналықтардың басым болуы жағына қарай езгереді, мүны аталық жыныстар особь-тарының тіршілік қабілеттілігінің шамалылығымен түсіндіруге болады.
1. ЖЫНЫСТАРДЫҢ ТАБИҒИ ЖАҒДАЙЛАРДАҒЫ АРАҚАТЫНАСЫ
Жыныстардың әр түплі организмдердегі арақатынасы ба-рынша ауытқып отырады. 100$: 0 & және 0$: 100 & -ге дейінгі кез келген арақатынастың дерлік мысалдарын табуға болады. Жыныстар арақатынасының өзгеруі онтогенез барысында әрекетететін факторларға, сондай-ақ генетикалық себептерге де бай-ланысты болуы мүмкін.
Соңғы жылдары бірқатар насекомдардан (дрозофиланың, қызыл қоңыздың кейбір түрлерінен) бір жынысты линиялар табылды. Бұл линиялардың үрғашылары тек қана аналық үр-пақ береді. Шағылыстыру кезінде бүл қасиет аналық линия бойынша түқым қуалайды. Бір жынысты линиялы шыбындар-дың гемолимфасында үсақ спирохета болатыны анықталған, сірә ол аталық зиготаны талғап зақымдайтын болса керек. Ата-лық типті зиготалардың қырылуыи үрықтанған жүмыртқалар-дың 50%-нің ждйылуына қарап анықтайды (10-тарауды қара-ңыз).
Жыныстардың арақатынасы генетикалық себептердің әсері-мен өзгеретін жағдайлар да белгілі. Мәселен, дрозофилада аутосомалардың бірінен гомозиготалы күйде аналық зиготалар-ды (2Х -)- 2Л) фенотиптік еркектерге айналдыратын рецессивті ген і табылған; бүл еркектер стерильді болады. і гені бойынша гомозиготалы ХҮ еркек особь өсімтал болып табылады. Егер қалыпты үрғашы (ХХі+ /+) көрсетілген ген бойынша (ХҮіі) гомозиготалы еркегімен шағылысатын болса, онда бірінші үр-пақта үрғашыларының ХХі+і, ал еркектерінің — ХҮі+і генотипі болады. Бүл шыбындардың ұрпағында үрғашыларының 'Д-нің гомозиготалы күйдегі і аллелі болады, олар фенотипі женінен €ркек особьтар болады. Бүл бірден-бір факт емес, мүндай қүбы-лыстар адам, жануарлар мен өсімдіктер үшін де сипатталған. Сүт қоректілер мен адамнан гомозиготалық күйі ХҮ генотипін-де аналық жыныс белгілерінің дамуына себепші болатын (Цт) гені сипатталған. Тестикулалық феминизация деп аталатын қүбылыстың мәні мынада: ХҮ генотипі бар особьтың қүрсақ қуысында тестикулалары, аталық бездері болады, бірақ қосым-ша жыныстық белгілерінің бәрі және мінез-қылығы — аналыққа тән болады. і\т генінің реттеуші ген, ал оның енімі (рецептор-лық белок) — реттеуші белок болып табылатынын зерттеулер көрсетті. Реттеуші белок нормада тестостерон гормонымен комплекс түзеді және осы арқылы аталық қосымша жыныстық белгілердің дамуына жауапты бірқатар қүрылымдық гендердің қызмет атқаруына жағдай жасайды. Мутанттық ген тестосте-ронмен әрекеттеспейтін бёлок түзіп шығарады, демек, особь-тың еркек ретіндегі дифференциациясы да бүзылады. Сүт қо-ректілер мен адамның аналық қосымша жыныстық белгілері индукцияланбайтын болып табылады және тканьдердің андро-генге реактивтілігі есебінен дамиды. Мысалы, жүгерінің бір үйлі есімдігін екі үйлі өсімдікке айналдыратын гендер табыл-ған. Рецессивті мутанттық ген 5і? ғомозиготалы күйде өсімдікті аталық өсімдікке айналдырады, өйткені ол тұқым бүрін жетіл-дірмей, тіршілігін жоюға себепші болады. Басқа мутанттық рецессивті ген (із) гомозиготалы күйде сыпыртқыда тұқым бу-рі мен түқымның дамуына себепші болады, ал бүл кезде тозаң-қаптар жетілмейді, яғни сыпыртқы аналық гүл шоғырына айналады. Қейде бір үйлі өсімдіктің аналық есімдікке айналуын цитоплазма анықтауы мүмкін (10-тарауды қара).
Табиғатта жыныстардың белгілі бір арақатынасын табиғи сүрыптау бақылап отырады, өйткені түрдің оптймальды санын өндіру мен түқым қуалайтын өзгергіштікті са.қтау үшін онын, зор маңызы бар.
2. ЖЫНЫСТАРДЫҢ АРАҚАТЫНАСЫН ҚОЛДАН РЕТТЕУ
Адам жыныстардың арақатынасын қолдан реттеуге тырыса-ды. Мүнда белгілі бір нәтижелерге, әсіресе жыныстық көбеюдін, жүйесіз типтеріне жағдай жасауда белгілі нәтижелерге қол жетті. Мәселен, Б. Л. Астауров жоғары температурамен әсер ету арқылы тұт кебелегінде партеногенез туғызды, мұнда мейоз болған жоқ, үрпақта 100% үрғашы особьтар алынды. Табиғи партеногенезде мейоз қалыпты өткен кезде ҮҮ особьтары қыры-латындықтан, тек қана еркек особьтар пайда болады. Андроге-незде бүкіл үрпақ тек қана аталық жынысты болады.
Электрофорездің жәрдемімек сперманы бөлу және бүдан кей-ін үрғашыларды «катодтық» ңемесе «анодтық» спермамен қол-дан үрықтандыру жолымен сүт коректілердегі (кролик, ит және т. б.) жыныстар арақатынасын қолдан реттеу жөніндегі жүмыстар көптеген жылдар бойы жүргізілді. Соңғы жылдары осы мақсатта еркектерді немесе ұрғашыларды бөЛінген сперма-мен иммунизациялау қолданылады. Қазірдің өзінде кеп үміт күттіретін нәтижелер алынды: топтарда бір жыныстың 80— 90%-ке дейінгі особьтары болады. Алайда бүл әдістер жануар-лардың үрықтанғыштығы мен өсімталдығын кенет кемітетін болғандықтан, әлі де практикада қолданылмай отыр.
Жынысты анықтаудан өзгеше, орта жағдайларының жыныс-тардың арақатынасын өзгертудегі ролі өте зор екендігіне және оның әсерін зерттеуге көптеген зерттеулер арналғандығына на-зар аудару керек. Ортаның әсерін мынаған келтіруге болады: жыныс хромосомаларының мейозда кездейсоқ ажырамауы, бел-гілі бір генотиптегі гаметалардың немесе зиготалардың талға-малы қырылуы, талғамалы үрықтануы, онтогенезде жыныстың қайта анықталуы. Әр түрлі орта жағдайлары жыныстар арака-тынасын 1 үрғашыға 1 еркек келетін өте дүрыс аракатынастан көп немесе аз дәрежеге дейін өзгерте алатыңын зерттеулер көрсетуде.Сонымеы, кейбір популяцияларда табиғи жағдайларда жы-ныстар арақатынасы 1: 1-ден өзгеше болуы мүмкін. Бүл шама-дан ауытқу генетикалық себептерге, сондай-ақ онтогенез бары-сында әрекет ететін факторларға байланысты болады. Объекті-нің биологиясы мен генетикасын ескерген жағдайларда жыныс-тар арақатынасын қолдан реттеуге болады.
VI бөлім
Онтогенездің генетикалық негіздері
Кез келген организмнің дамуы — оның клеткалары мен тканьдерінің тек авторепродукциясы ғана емес, заңды өзгеріс-тердің ұзын тізбегі, химиялық, физикалық, морфологиялык жә-не функционалдық тұрғыдан жаңаның пайда болуы. Массасы не бары 0,0015 мг шамасында болатын адамның жұмыртқа клеткасының ұрықтанғаннан кейін массасы 3 кг шамасында болатын баланың дамитынын ескерсек жеткілікті. Жылдар өтеді, адам өзін қоршаған дүниеңі талдауға және оны өзгерту-ге кабілетті бола бастайды.
«Онтогенез» деген үғымға ұрықтанудан немесе жұмыртқа-ның дамуын активтендіруден басталатын және организмнің өлімімен аяқталатын даму процесі кіреді. Жүмыртқа клетка-сында да, сперматозоидта да дайын белгілер болмайды, оларда сыртқы және ішкі ортаиың белгілі бір жағдайларында іске асырылатын көп клеткалы организм дамуының программа-сы ғана салынған.
Жеке даму генотип системасымен анықталады, онда гендер әсерінің ерекшелігі, уақыты, орын реттілігі проғраммаланған.
Онтогенездің түқым қуалайтын негіздерін зерттейтін генети-каның бөлімі феногенетика немесе онтогенетика деп аталады.
21-тарау. ДИФФЕРЕНЦИРОВКАНЫҢ (ЖІКТЕЛУДІҢ) ГЕ-НЕТИКАЛЫҚ НЕГІЗДЕРІ
Қөп клеткалы организм есуіыің және жеке дамуының негі-зінде клеткалардың митоздық бөлінуІ жатады. Митоз — тең тү-қым қуалайтын бәліну, осыған байланысты организмнің ма-манданған әр түрлі тканьдеріндегі (мидың, бүлшық еттердің, терінің, бауырдың т. б.) клеткалардық генотиптері үқсас бола-ды. Бүл жағдайда мынадай сүрақ қойылады: онтогенездің ба-рысында клеткалар мен тканьдері дифференцировкасының гене-тикалық механизмдері қандай болуы мүмкін?
Онтогенезді генетикалық зерттеудің бастапқы кезеңі: бір ген — бір белгі, немесе ген (ДНК) -РНК — белок —... — белгі принципіне сәйкес белгіні қалыптастырудағы геннің әсеріне анализ жасау болып табылады. Жеке дамудың тұқым қуалау-шылық негіздерін зерттеудегі басты проблемалар: гендердің дифференциалды активтілігін зерттеу және ген — белгі. тізбегін-дегі аралық буындарды анықтау болып табылады.
Жануарлар мен өсімдіктердің онтогенезінде бірқатар негіз-гі процестер: өсу, тканьдердід дифференцировкасы, морфогенез, яғни органдар мен белгілердің қалыптасуы жүзеге асырылады.
1. АЛҒАШҚЫ ДИФФЕРЕНЦИРОВКА
Үрықтанғанға дейінгі жұмыртқаның дифференцировкасы.
Жануарларда алғашқы морфологиялық дифференцировка жұ-мыртқа цитоплазмасының қүрылымымен және оның беткі қа-батымен (кортекспен) қамтамасыз етілетіні белгілі. Мысалы, кейбір амфибийлермен омыртқасыздардың ядросыз жүмыртқа-лары оларды активтендіргеннен кейін бластула стадиясына де-йін даму қабілетін сақтайды. Кортекс функционалдық дискрет-ті. Оның эктодерма түзілетін анимальды зонасы, мезодерманың негізі қаланатын «сүр орақ» зонасы (гаструляция осы арадан басталады) және энтодерма түзетін вегетативті зонасы болады. Сонымен, тіпті үрықтанғанға дейін жұмыртқа дифференциялан-ған болады. Ұрықтанғаннан кейін үрықтың ерте стадияларын-дағы дамуын анықтайтын жүмыртқаның дифференциялануы бүдан да гері нәзік болады.
Митоздық бәліну жолымен пайда болған бластомерлерде бірдей геномдар болады, бірақ оларда кортекс және цитоплаз-
| Анимальдь/к, полңіс |
ма учаекелерінің көлемі бір-дей болмайды (86-сурет).
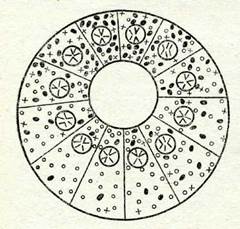 |
| 86-сурет. Жұмыртқаның беліну пр |
| цесінде цитоплазмалық құрылымдардын, ажырауы біркелкі болмайтынды- |
| ғын көрсететін схема. Түрлі цитоплазмалық құрылымдар крест, нүкте, деқгелекпен белгіленген. |
| Вегетативтің. полюс |
Осының нәтижесінде түрлі бласто-мерлердегі әр түрлі гендердің саналуын реттеуші қызметін атқаратын, сөйтіп дифферен-цировканың барысына әсер етіп, оларда әр түрлі болып құралғак цитоплазма болады. Ядро мен цитоплазма диффе-ренцировка процесінде өзара байланыста болады, осыған орай жүмыртқаның цитоплаз-масы мен қортекс қабатыиын, преформациялануы аналық организм генотипінің қызметі-нің ыәтижесі болып табылады. 10-тарауда үрпақтың белгі-лерінің дамуына әсер етуші іштоплазманың ерекшеліктерін генетикалық анықтауға мысал-дар келтірілген болатын, мы-салы, үлу қабыршағының солға және оңға қарай бүра-лушылығы (58-суретті қара). Жануарлар мен өсімдіктерде кен, тараған партеногенез фактісі шешесінің генотипі өз беті мен ұрпақтың қалыпты өзін-өзі өндіруін қамтамасыз ете ала-тындығына дәлел бола алады.
Шешесінің генотипі әдетте жұмыртқа клеткасының генотипі-мен сәйкес келмейтінін есте ұстау керек. Жұмыртқа клеткасы-ның қалыптасуына диплоидты аналық организмнің барлық ген-дерінің жиынтығы қатысады. Мейоздан кейін онда гендердің гаплоидты жиынтығы сақталады. Алайда цитоплазмада дипло-идты аналық организмде түзілген және богенез құрылымында қалыптасқан гендердің өнімдері қалады. Аналық организм ал-дын ала әзірлеген информация ретінде, міне осылар жұмыртқа-ның бастапқы даму фазаларын қамтамасыз етеді, атап айтқан-да ұрықтың дамуы барысында белоктарды синтездеу үшін мат-рица қызметін атқаруға қабілетті информациялық РНК күйінде болады. Мысалы, шпорц бақаның (Хепориз 1ае\гіз) үрығында соңғы бластулаға дейін «лампа шеткалары» стадиясында ооцит синтездеген тек қана и-РНҚ болады және өз қызметін атқара-ды. Соңғы бластулада ғана үрық геномының активтендірілуі есебінен и-РИК-ның синтезделуі басталады. Онан басқа, клетканың цитоплазмалық органоидтарында ядро сияқты информацияны сақтауға қабілетті, өздерінің нуклеин қышқылдары — ДНК және РНК-сы болады.
Бөлшектену кезіндегі дифференцировка. Ұрықтанғаннан кейін аталық организмнен алынған гендердің де қызметі керінетін онтогенездің нағыз өзі басталады.
Егер гендер бүкіл онтогенезді, организмнің барлық белгіле-рі мен реакциясын бақылайтын болса, онда кептеген сүрақтар туады: дамудың түрлі сатыларында гендердің барлығы бір мезгілде әсер ете ме нед/гесе тек кейбіреулері ғана ма? Олардың іс-ке қосылғаны немен анықталады? Гендердің ерекше әсері қа-лайша жүзеге асырылып отырады?
Бұл сүрақтардың кейбіреулеріне ядроларды транспланта-циялау тәжірибелері жауап беруге мүмкіндік береді. Бақаның (Яшга езсиіепіа) үрықтанбаған жүмыртқа клеткасын укол са-лу арқылы активтендіреді және оның ядросын алып тастайды. Бүдан кейін оған бластула, гаструла және с. с. стадиядағы үрық клеткасының ядросын алып орнатады. Егер донор-клетканың ядросы дифференцияланып кеткен болса, онда оны ауыстырып отырғызғаннан кейін жұмыртқа — реципиент қалыпты үрық бер-мейді. Егер донордың ядросында әлі дифференция болмаса және толық дамуды қамтамасыз ететін қабілеттілігі сақталса, он-да жүмыртқа клетка—реципиент, тіпті ит балық болып қалыптасқанға дейін, қалыпты бөлшектене береді. Егер ядро донордың бластула немесе ерте гаструла стадиясындағы клеткасынан алынғаи болса, онда жүмыртқа-реципиенттен қалыпты ит балық дамитыны айқындалды. Демек, дамудың ерте стадияла-рындағы клеткалардың ядролары әлі дифференцияланбаған жә-не ол зиготаның ядросымен тең бағалы болады. Соңғы ғастру-ла клеткаларынан ауыстырылып отырғызылған ядросы бар жүмыртқадан әдетте ұрық дамымайды; демек, гаструляцияның басталу кезеңінде қайталанбайтын дифференцировкасы жүреді. Алайда, кейбір ерекше жағдайларда осындай дифференциялан-ған клеткалардың ядроларын, мысалы ит балық ішегінің клет-каларын ядросыз жүмыртқа клеткасына ауыстырылып отырғы-зылғанда урықтьщ қалыпты дамуын, метаморфозды және тіпті ұрпақ бере алатын бақаның дамуын қамтамасыз ететіндей қа-білет болады.
Тканьдер арасындағы индукциялық қатынастар. Даму ба-рысында гаструладан бастап тканьдер арасында индукциялық қатынастар, яғни бір ткань екінші тканьнің даму сипатына ба-ғыт бере отырып әсер етеді. Мысалы, омыртқалыларда гастру-ляцияның барысында хорданың бастамалары эктодерманың бел-гілі бір ауданымен түйіседі, соның нәтижесінде эпидермалық клеткалар барлық эктодерма сияқты терінің эпителийлеріне емес, нерв системасына дифференцияланады. Индукция меха-низмі мынадай: тканьнің клеткаларында арнаулы заттар «ин-дуктор» түзіледі, ол көрші индукцияланатын тканьге ауысып, оның даму жолын өзгертеді. Анығында, хорда бастамасы клеткаларының гендері қызметінің өнімдері эктодерма клеткаларын-дағы нерв системасының дамуын анықтайтын сондай гендердің орекетін активтендіреді.
2. ДИФФЕРЕНЦИРОВКАНЫҢ ГЕНЕТИКАЛЫҚ МЕХАНИЗМДЕРІ
Гендердің дифференциалды активтілігі. Көп клеткалы орга-низмнің барлық клеткаларында гендердің үқсас жиынтықтары болады, бірақ түрлі уақытта әр түрлі тканьдерде сан алуан ген-дер әрекет етеді, солардың арқасында дифференцировка іске асады. Ген әсерінің реттелуі әр түрлі: репликация, транскрип-ция және трансляция деңгейлерінде іске асады.
Репликация деңгейіндегі реттелу. Реттелудің бүл типі дәл осы мезгілде қажетті немесе қажетсіз гендер көшірмелерінің санын көбейту немесе азайту есебінен жүзеге асырылып оты-рады.1
Кейбір жағдайларда гендердің саны емес, хромосомалардың немесе ДНК молекулаларының саны көбейеді. Дифференциров-ка процесінің барысында интенсивті қызмет етіп жатқан соматикалық клеткаларда полиплоидизацияның жүруі мүмкіні Егер көп клеткалыларда полиплоидты ядролар тіпті кездеспеуге жа-қын болса, қарапайымдарда олар ірі системалық категориялар-ға тән болады. Мәселен, инфузорияларда ядрольщ дуализм — бір мезгілде қатарынан екі ядроның — диплоидты генеративтік [микронуклеус) және вегетативтік немесе соматикалық (макро-нуклеус) ядролардың кездесуі орын алады. Қейбір инфузория-лардың макронуклеусі жоғары полиплоидты келеді.
Ядролық дуализм — түқым қуалайтын информацияны үр-пақтан үрпаққа беру үшін жауапты диплоидты микронуклеус (оның гендері фенотип бойынша көрінбейді және активтелмегён жағдайда болады) пен инфузорияның бүкіл тіршілік әрекетін бақылайтын жоғары полиплоидты (10000—13000 п) макро-нуклеус арасындағы өзінше «еңбек бөлісу». Плоидтың жоғары дәрежеде болуының арқасында макронуклеустің метаболиттік активтілігі де жоғары болады, бұл активтілік гендер өнімдерін көптеп шығару арқылы қамтамасыз етіледі.
Инфузорияның онтогенезінде макронуклеус өзінше ерекше дифференцияға ұшырайды. Коньюгацияның алдында ол ыды-райды, ал коньюгация аяқталғаннан кейін бөлінген диплоидты синшрионнан (мейозға ұшыраған екі гаплоидты микронуклеус-тің қосылу өнімі) қайтадан түзіледі. Бұдан кейін эндомитоздық бөлінудің арқасында макронуклеустің плоидтылығы инфузорияның осы түріне тән мөлшерге дейін көбейе береді.
Макронуклеустің әрбір гаплоидты жиынтығының барлық хромосомалары шеткі жақтарымен тұрақты коньюгация жағдайында болады және әрбір мұндай жиынтық тізбек тәрізденіп шұбатылып жатады. Инфуз-ориянын, жыныссыз көбейетіндігіне қарамастан макронуклеус амитоздық жолмен бөлінеді, хромо-сомалар коныогациясының ерекше сипаты және оның жоғары плоидтылығы, жас ядроларға бірнеше хромосом жиынтықтары-ның түсуін қамтамасыз етеді, ол жиынтықтардың саны одан әрі эндомитоздық бөліну аркасында осы түрге тән мөлшерге дейін көбейе береді. Осы себептер бойынша макронуклеус регенерациялануға қабілетті болады, бір клеткалы организм үшін бұл аса маңызды.
. Соңғы жылдары кейбір инфузориялардың гендер көшірмеле-рі санының кебеюіне басқа да жол — политенизация ашылды. Онан соң политенді хромосома жеке хромомерлерге, тіпті ген-дерге ыдырайды, олар макронуклеус бөлінген кезде жас ядро-ларға кездейсоқ таралады.
Жануарлар мен есімдіктердің кейбір түрлерінің дифферен-цияланған тканьдерінің клеткаларында эндомитоз және политения қүбылыстары байқалады. Мәселен, кейбір өсімдіктердің крахмал түзетін клеткаларында, хар түқымдас балдырлардың маманданған клеткаларында, сүт қоректілердің бауыры мен ішек эпителийінде эндомитоз жүреді, соның нәтижесінде бүлар-да полиплоидия орын алады. Хромосомалардың политендік жағдайы сілекей бездерінің интенсивті қызмет жасап жаткан ядроларына, ішекке және қос қанаттылар личинкаларынын, мальпигий түтікшелеріне тән болады. Осы мысалдардың бәрі хро-мосомалардың күйі мен санының организмнің соматикалық клеткаларының морфофизиологиялық дифференцировкасымен сез-сіз байланысы бар екенін көрсетеді.
Жекелеген гендердін, кебею қүбылысы — гендердің ампли-фикациясы анағүрлым кең таралған. Мәселен, кейбір омыртқа-лылардың ооциттерінде «лампа щеткалары» типтес хромосома-лар табылды (3-тарауды қара). «Лампа щеткаларының» ілмек-тері хромосоманың деспиралданған учаскелері болып табылады. Олардан и-РНК-ның көп мөлшері табылды, демек гендер ак-тивті түрде қызмет етеді. Каллан әрбір иректі бір геннің көптеген репликтерінің линиялы жиынтығы деп санайды, осынын, арқасында и-РНК-ның көптеген санын синтездеу мүмкіндігі арта түседі, оны соңынан эмбрион пайдаланады (225-бетті қа-ра). Гендердің амплификациясы кеңінен таралған құбылыс, ол өсімдіктердіқ, жануарлардың және адамнын, қызмет атқарып жатқан әр түрлі клеткаларынан табылды. Мүндай гендер қыз-мет атқармайтын жерде, олар хромосоманың гетерохроматиндік аудандары болады (2-тарауды қараңыз).
Транскрипция деңгейіндегі реттелу. Бұл механизм транск-рипция процесінің түрлі сатыларында жүмыс істей алады. Мә-селен, РНҚ-полимеразаның жекелеген гендерге туыстығы өзгеруі мүмкін және осының нәтижесінде осы гендерден транскрип-цияланатын и -РНК-ның саны өзгереді. Санаудың басы немесе соңы өзгеруі мүмкін, осының нәтижесінде қысқарған немесе үзарған и -РНҚ-нық молекулалары пайда болады. Және ең соңында метаболизмнің ерекшеліктеріне байланысты геномның жекелеген учаскелері РНК-полимеразаны өткізуі немесе өткіз-беуі мүмкін. Бұл оперондыщ реттелу деп аталады. Уақыт жағы-нан гендер активтілігі реттелуінің осы системасын бактериялар-да Ф. Жакоб пен Ж. Моно ашты және ол гендердің үш типінең: белок-ферменттерді синтездеуге жауапты бірнеше структуралық гендерден, ген-оператордан, ген-реттеушіден түрады. Ген-опера-тор және онымен тығыз тіркескен структуралык, гендер оперон қүрайды (87-сурет).
Кызметі жағынан бір-біріне байланысты ферменттерді ты-ғыз тіркескен және и -РНК-ның бір молекуласын транскрип-циялайтын структуралық гендер кодтайды? Осы гендер транск-рипцияның реттелуші бірлігі — оперонға кіреді. Структуралық гендерден транскрипцияның болуы немесе болмауы ген-опера-тормен анықталады. Оператор көбінесе РНК-полимеразаның қосылу нүктесі—промотормен — структуралы гендер аралығын-да жатады, яғни олармен тығыз тіркескен. Егер оператор бос
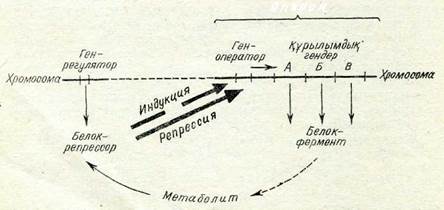 |
| 87-сурет. Реттеуішоперон системасының жұмысын көрсететін схема. (Түсініктеме тексте беріліи отыр). |
болса, РНК- полимераза одан өтіп кетіп, и -РНК-ны синтездеуге кіріседі. Егер оператор ерекше белокрепрессордың бо-луына байланысты бос болмаса, и -РНК синтезделмейді. Белок-репрессор оперонмен тығыз тіркеспеген ген-реттеушімен кодта-лады. Репрессордың өз операторынан орын алуы қабілеті эффекторлармен — физиологиялық сигналдар болып табыла-тын кішіректеу молекулалармен анықталады. Олардың іс-әре-кет сипаты метаболизмнің қандай сатысын бақылайтынына байланысты болады. Мәселен, клеткада реттелмейтін әр уақытта болатын және синтезделуі конститутивтік ферменттер бар.
Ферменттердің басқа тобы оперон системасымен реттеледі. Бұл, біріншіден, катаболизмге қатысатын индуцибелъдік фер-менттер. Олардың синтезі осы ферменттердің әрекеті мен баға-лы сапасын жоятын заттардың клеткада жиналуы кезінде басталады, яғни бұл зат өз оперонының белок-репрессорымен қо-сылатын индуктордың немесе эффектордың қызметін атқарады, ояы оператордан бөлінуге мәжбүр етеді, сөйтіп транскрипцияны іске қосады: оперон жұмыс істейді, белок синтезделеді. Бұл индукция жағдайы.
Екінші жағдай анаболизмге қатысатын (биосинтез) репрес-сибелъді ферменттерге қатысты. Бұл жағдайда клеткаларда метаболиттің — осы биосинтез тізбегінің ең соңғы өнімі болып табылатын эффектор жинақталады, ол ферменттердің одан әрі синтезделуін тоқтатады. Метаболит-эффектор репрессорға қосы-лып оны оператордан орын алуға мәжбүр етеді, сөйтіп струк-туралық гендерден транскрипциялану жабылады, ферменттер-дің синтезделуі тоқталады. Бұл репрессия жағдайы.
Хромосомалардың қайта құрылуы нәтижесінде структуралық гендердің ген-оператордан бөлінуі бұл гендерді бастапқы оператордың бақылауынан шығарып жіберуі және оларды бас-қа оперонмен байланыстыруы мүмкін. Мәселен, ішек таяқша-сында делеция алынды, ол делеция лактоздық және пуриндік оперондардың фрагменттерін қосты, осы күрделі оперонның операторы пуриндік, ал структуралық гендері — лактоздык бол-ды. Нәтижесінде ортаға пуринді қосқанда лактоз облысында белоктардың күшті репрессиясы жүрді. Хромосомалық қайта қүрылу нәтижесінде оператор мен әр түрлі оперонға жататын структуралық гендердің қосылуы ген орналасуы әсерінің моле-кулалық механизмі бола алады (13-тарауды қараңыз).
Ген-реттеушілердің жекелеген мысалдары, мысалы, адамда тестикулярлық феминизациялау генінің болатыны белгілі бол-ғанымен, кәп клеткалы организмдерде гендер активтілігі ретте-луінің оперон системасы табылған жоқ. Бүларда ген активтілігін басқаратын системаның жүмыс принципі басқаша деп топ-шылауға болады, өйткені бактерияға қарағанда жоғары эукариоттардың клеткаларындағы гендердің басым көпшілігі (80—95%-ке дейіні) қызмет атқармайды, яғни репрессия жағ-дайында болады.
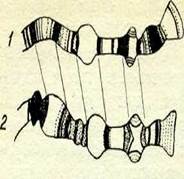 |
| 3-сурет. Қалыпты даму кезде |
Дрозофила личинкаларыньщ сі-лекей бездерінің клеткаларындағы гигант хромосомалардың белгілі бір дискілерін байқау "үстінде зерттеушілер хромосомалардың белгілі бір жерінде личинкалар дамуының
дрозофиланьщ алып хромосо- белгілі бір стадиясында кейбір дисма

