 2015-07-14
2015-07-14 1753
1753Этот же феномен использовался для объяснения ряда эффектов обусловливания в другой модели (Л. Л. Воронин, 1982). Синаптические аналоги условного рефлекса создавались в зоне коркового представительства передней конечности бодрствующего кролика сочетаниями электрического раздражения двух точек коры. Условным стимулом служило подпороговое (неэффективное) раздражение коры на некотором расстоянии от исследуемого коркового нейрона, а безусловным — пороговое для постсинаптической реакции раздражение поверхности коры над исследуемым корковым нейроном. При активирующих формах условной связи в ответ на условный стимул возникали ВПСП, амплитуда которых постепенно возрастала до критического уровня генерации потенциалов действия (рис. 53). При отмене подкрепления, то есть при угашении, амплитуда ВПСП постепенно уменьшалась с прекращением генерации спайковых разрядов.
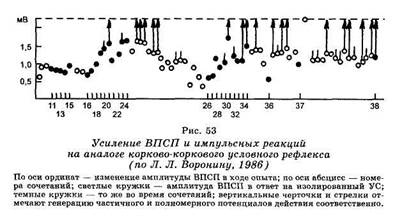
При тормозных формах временной связи условный стимул постепенно вызывал развивающееся угнетение механизма генерации импульсных разрядов, которое не сопровождалось существенной гиперполяризацией мембраны клетки или достоверным уменьшением ВПСП. При этом установлено, что дифференцирование одним и тем же нейроном подкрепляемых и неподкрепляемых стимулов реализуется различными синаптическими входами и поддерживается благодаря механизмам гетероси-наптического облегчения.
Одной из моделей аналога временной связи на микропопуляции нейронов коры больших полушарий является микроионофоретическая аппликация медиаторов (глутамата и ацетилхолина), используемых в качестве аналогов ассоциируемых раздражителей (Б. И. Котляр, 1986). Установлено, что отдельные нейроны микропопуляции в радиусе действия медиаторов обнаруживают специфические ассоциативные перестройки своей активности. Последние могут заключаться в следующем: изменения реактивности к сигнальному раздражителю, воспроизведение реакции на подкрепляющий раздражитель в ответ на действие условного сигнала и реакция на время в границах действия ассоциируемых раздражителей.
В. С. Русинов (1969) пришел к выводу, что суммаци-онный рефлекс и доминанта представляют собой начальные стадии образования любого условного рефлекса и сами по себе могут служить удобной моделью для изучения простых форм временной связи. Им была разработана модель корковой поляризационной доминанты, состоящая в том, что поляризация постоянным током участков сенсомоторной коры кролика при действии ранее неэффективных раздражителей — световых вспышек, приводила теперь к ритмическому сокращению передней конечности (корковая проекция которой поляризовалась) в такт световой стимуляции. Установившаяся под влиянием поляризации новая форма реагирования на раздражитель сохраняется до 30 мин после выключения тока.
Эта модель послужила существенным толчком для выявления пластических характеристик корковых нейронов. Установлено, что поляризация слабым постоянным током увеличивает активность полисенсорных ней-
ронов, что является одним из непременных механизмов возникновения движения на ранее индифферентный раздражитель. Причем функциональные и морфологические сдвиги происходят не только в пирамидных нейронах, но и в интернейронах и глиальных клетках, которые также подвергаются поляризации.
Все основные свойства доминанты: повышенная возбудимость, стойкость возбуждения, способность к суммированию возбуждений, инерции, торможению и рас-тормаживанию — были обнаружены на вышеописанной модели. В доминантном очаге происходит установка на определенный уровень стационарного возбуждения, способствующий суммированию ранее подпороговых возбуждений и переводу на оптимальный для данных условий ритм работы, когда этот очаг становится наиболее отзывчивым и тормозятся другие работающие очаги. Иными словами, образуется единый функционально организованный ансамбль или система. Подчеркивается особая роль нейронов нижних слоев коры, которые дольше сохраняют ритм раздражения, чем клетки верхних слоев. Обращается серьезное внимание на существование в системе базальных дендритов пирамидных клеток дендро-дендритических контактов, которые рассматриваются как важнейший субстрат межнейронных отношений. В. С. Русинов полагает, что наиболее адекватным отражением функционального состояния системы нейронов является уровень их постоянного потенциала, а следовательно, элек» тротонические влияния имеют наиболее существенное значение для образования временной связи. Поляризационный очаг, созданный постоянным током, служит удачной моделью для изучения поляризационно-электрото-нической гипотезы формирования доминанты и простых форм временных связей (В. С. Русинов, 1969).
Специальное внимание к нейроглиальным взаимоотношениям при формировании клеточных аналогов условного рефлекса привлек А. И. Ройтбак (1969). Он выдвинул глиалъную гипотезу, согласно которой в основе образования и укрепления временных связей лежит мие-линизация центральных аксонов, возбуждающихся от условного раздражения. При действии безусловного раздражения в соответствующих областях коры происходит
деполяризация глиальных клеток: возбуждение большого числа нервных элементов импульсами из специфических и неспецифических путей приводит к выделению в межклеточные щели и аккумуляции ионов калия. Автор полагает, что деполяризация олигоденд-роцитов служит сигналом к образованию миелина; условием миелинизации является совпадение во времени деполяризации олигодендроцитов и физико-химических изменений в прилегающих немиелинизированных пре-синаптических терминалях после их предшествующего возбуждения (рис. 54).
В последние годы широко развернулись исследования механизмов временных связей на животных с относительно простой нервной системой. Наиболее удобными для экспериментирования оказались моллюски. Простейшей моделью служил полностью изолированный нейрон висцерального ганглия виноградной улитки. Локальная аппликация ацетилхолина сочеталась с внутриклеточной инъекцией деполяризующего тока, в результате было показано, что изолированные нейроны моллюсков (то есть лишенные своих сетевых свойств) не способны формировать ассоциативный ответ в виде воспроизведения безусловной реакции сигнальным раздражителем.
Клеточные механизмы образования условного рефлекса на морском моллюске аплизии изучались Э. Кенделом (1980). Исследовался оборонительный рефлекс: условный
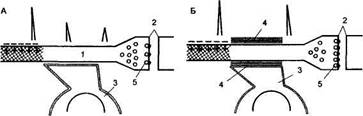
Рис. 54
Схема процесса образования временной связи (по А. И. Ройтбаку, 1972)
А — «потенциальный» синапс; Б — образование временной связи (превращение «потенциального» синапса в «актуальный») 1 — пресинаптический участок нервного окончания; 2 — синаптическая щель; 3 — олигодендроцит; 4 — миелиновая оболочка; 5 — медиатор в синаптических пузырьках. Эффективность проведения тока по терминальному волокну увеличивается вследствие формирования вокруг него оболочки из олигодендроцита.
сигнал — умеренной силы раздражение сифона, вызывающее втягивание жабры в мантийную полость, безусловный раздражитель — сильная стимуляция хвоста. В результате сочетаний защитная реакция на стимуляцию сифона усиливалась. Удобство этой модели состояло в том, что был точно определен нейронный состав рефлекторной дуги, в которую входят 24 сенсорных нейрона, образующие прямые связи с интернейронами и 6 моторными нейронами, иннервирующими жабру и сифон. Стимуляция хвоста возбуждает нейроны, контактирующие с пресинап-тическими окончаниями сенсорных нейронов.
В данной модели обнаружились явления сенситиза-ции, привыкания и нейрогуморального (субклеточно-молекулярного) воздействия безусловного раздражителя на всех этапах выработки временной связи. Суммация заключалась в неспецифическом гетеросинаптическом облегчающем эффекте — увеличении ВПСП, связанном с увеличением выброса медиатора из пресинаптического окончания. Суммация этих процессов приводит к возникновению длительной деполяризации, составляющей сущность проторения путей при образовании временной связи. Процессы привыкания связаны с ограничением потоков кальция в пресинаптические окончания, что приводит к гиперполяризации нейронов и снижению их ответа на безусловный раздражитель. Нейрогуморальный эффект безусловного раздражителя достигался благодаря механизмам регуляторных нейропептидов (см. гл. 8).
Э. Кендел (1980) считает, что в основе условной реакции втягивания жабры у аплизии лежит механизм пресинаптического облегчения (сенситизации), который формируется при конвергенции раздражителей, приводящих к спайковой активности нейрона, и активации этого же нейрона через модулирующий вход от безусловного раздражителя. Аналогичный механизм допускается и у позвоночных животных, хотя прямые доказательства этого отсутствуют.
Какими бы информативными для задач общей нейрофизиологии ни были аналитические подходы к исследованию наркотизированных или обездвиженных препаратов позвоночных и беспозвоночных животных с использованием различных моделей и аналогов условных
рефлексов, их конечной целью является расшифровка тех феноменов мультинейронной активности, которые регистрируются на бодрствующем мозге при выполнении животным того или иного целостного условнорефлекторного акта. Сегодня мы еще не можем использовать весь арсенал сведений аналитической нейрофизиологии для понимания клеточной активности бодрствующего мозга. Различия двух основных подходов базируются на степени проявления свойства пластичности, а именно — высокой изменчивости реакций одного и того же нейрона у бодрствующего животного и значительно большей стабильности реагирования нервных клеток у наркотизированных препаратов.
§ 35. НЕЙРОННАЯ ОРГАНИЗАЦИЯ УСЛОВНОРЕФЛЕКТОРНОГО ПРОЦЕССА
Данные о перестройке нейрональной активности при выработке условных рефлексов у бодрствующих животных представляют существенное значение для понимания функциональной организации целостной системы обучения. Такие изменения реакций нейронов могут проявляться практически во всех исследованных структурах мозга. Логично допустить поэтому, что возможность образования ассоциаций является базовым свойством нервной системы.
В связи с тем, что сигнальная сущность условного рефлекса состоит во временном сочетании, то есть в ассоциации раздражений, представлялось необходимым, в первую очередь, выделить перестройки нейрональной активности, связанные с ассоциацией, в отличие от всех остальных сопровождающих изменений в активности нейронов.
Р. Томпсон рекомендует для этой цели использовать три критерия: 1) специфичность изменений активности на условный сигнал в сравнении с изменениями, вызываемыми индифферентными раздражителями; 2) параллельность развития изменений нейронной активности и компонентов поведенческого условного ответа и 3) градуальный рост выраженности пластических изменений
на нейронном уровне в процессе выработки поведенческого навыка.
Другие авторы (Э. Кендел, 1980) предложили для той же цели рассматривать два независимых критерия: 1 — природа ответа на условный сигнал, то есть является ли условный ответ новым (классический условный ответ) или модификацией условной реакции на сигнальный раздражитель (альфа-условный ответ), и 2 — специфичность реакции для последовательного сочетания раздражителей. Иными словами, не может ли аналогичный ответ развиваться при случайном предъявлении тех же раздражителей (при процедуре псевдообусловливания) или определяться сенситизацией.
Б. И. Котляром (1986) предложена классификация перестроек активности нейронов в процессе условнореф-лекторной деятельности (табл. 5). Принципиальным является введение в состав условнорефлекторного процесса тонического типа активности нейронов. Основанием этому служили специальные исследования с переключением разнородных условных рефлексов, которые позволили рассматривать центральный мозговой тонус в качестве непременного компонента процессов обучения и памяти.
Таблица 5
Классификация разных видов функциональных перестроек
активности нейронов головного мозга при формировании
и реализации временной связи (Б. И. Котляр, 1986)
| Ассоциативные перестройки | Неассоциативные перестройки | ||
| физического типа | тонического типа | физического типа | тонического типа |
| Перестройки, отражающие уровень центрального тонуса, характерного для реализации временной связи | Перестройки, характерные для тонической составляющей ассоциативного процесса | Перестройки, отражающие динамику ориентировочной реакции | Перестройки, отражающие динамику ориентировочной реакции |
| Специфические перестройки, воспроизводящие безусловный ответ | Перестройки, связанные с доминирующей мотивацией | Перестройки, связанные с доминирующей мотивацией | |
| Перестройки, отражающие реакции на время в рамках действия ассоциируемых раздражителей | |||
| Перестройки, связанные с условной ориентировочной реакцией |
¥ I
По результатам обучения на уровне отдельной клетки можно судить, что наиболее существенные изменения характерны для корковых полисенсорных нейронов (М. Я. Рабинович, 1975), функциональные свойства которых по мере сочетаний претерпевают эволюцию, отражающую становление его афферентной специализации. Так, до первых сочетаний стимула с подкрепляющим рефлексом нейроны проявляют преимущественно моносенсорные реакции, то есть большинство из них отвечает на стимулы адекватной сенсорной модальности стабильными коротколатентными разрядами. По мере последующих сочетаний эти же нейроны приобретают способность отвечать на стимулы различных сенсорных модальностей и параметров; их активность становится полисенсорной. Этот период соответствует фазе генерализации условного рефлекса, когда положительное действие обнаруживают не только сигнальный, но и другие сенсорные раздражители. Вероятно, именно в этот период данная рефлекторная система начинает приобретать доминантные свойства, выделяться среди прочих и подкрепляться разнообразными воздействиями. Наконец при достаточно упроченном условном рефлексе ответы нейронов снова приобретают моносенсорный характер, но теперь уже это специальные моносенсорные реакции на стимул определенной модальности и физических параметров, то есть избирательная реакция только на тот стимул, который приобрел сигнальное значение для поведенческого акта.
Следует отметить, что характеристики процесса привыкания реакций нейронов при монотонных предъявлениях сенсорных стимулов не коррелируют с процессом ассоциативного обучения. Одни нейроны в популяции могут проявлять феномен привыкания, а другие — сен-ситизацию, следовательно, нейрофизиологические механизмы привыкания и ассоциативного обучения могут быть различны по своей организации. Если в основе привыкания лежит уменьшение числа квантов медиатора, ведущее к снижению амплитуды постсинаптических потенциалов, ассоциативное обучение может быть связано со сдвигом эндогенной активности обучаемого нейрона. Однако нельзя исключить возможность причинно-след-
279
ственнои связи между привыканием и сенситизацией. Действительно, на одном и том же мультисенсорном нейроне можно наблюдать в процессе предъявления стимулов оба явления сразу. Например, в начале процедуры привыкания ответа на световой стимул, электрокожный стимул, подаваемый в интервале функциональных сдвигов после светового стимула, оказался неэффективным. Однако по мере ослабления ответной реакции нейрона на световой стимул (привыкание) начинает появляться ответ того же нейрона на электрокожный стимул (сенситизация). Последний достигает своего максимума при отсутствии ответа нейрона на световую вспышку (А. С. Батуев, 1978).
Преобладание того или иного механизма существенно зависит от сигнальной значимости предъявляемого раздражения. Так, в опытах на бодрствующих обезьянах было установлено, что сенсибилизация в реагировании нейронов лобной коры возникает лишь в том случае, если раздражители прямо или косвенно связаны с мотивационной сферой животного, сигнализируют об опасности или несут на себе элементы новизны. Привыкание ответов тех же нейронов наблюдалось в тех случаях, когда используемые раздражители были индифферентными или хорошо знакомыми для животного. Однако если ранее индифферентный агент приобретал качества сигнального фактора, будучи сочетай во времени с биологически значимым болевым агентом, то картина резко менялась (рис. 55). Ранее угашенные нейронные ответы уже через 9-10 сочетаний возобновляются, что характеризуется всеми чертами выработки условного рефлекса и подтверждается параллельной регистрацией ЭМГ рабочей конечности. Такой пластический характер муль-тисенсорной конвергенции к нейронам лобной коры, диапазон и устойчивость которой определяется сигнальной значимостью раздражителя, в конечном счете формируется доминирующей мотивацией и системой выработанных ранее временных связей.
По критерию наличия или отсутствия изменений реакций нейронов нельзя считать, что обучение характерно лишь для определенных отделов или уровней мозга. Имея в виду неравноценность отделов мозга для
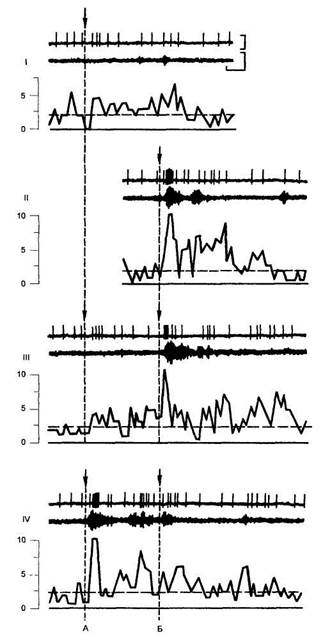
Рис. 55
Динамика импульсной активности коркового нейрона лобной области у обезьян и электромиограмма (ЭМГ) мышц плеча при
выработке условного рефлекса (по А. С. Батуеву и др., 1981) I — (сверху вниз): нейронная активность, ЭМГ и график изменения средней частоты (по девяти предъявлениям) при световом раздражении; // — при кожном раздражении (уколе); III и IV — при сочетании светового раздражения с уколом соответственно при 1-9-м и 10-18-м их сочетаниях; V — график изменения латентного периода ЭМГ сдвигов. На графиках I-IV: по оси абсцисс — время в с; по оси ординат — средняя частота в имп/с при At — 100 мс; горизонтальной штриховой линией указан уровень фоновой активности, на графике V — по оси ординат -- порядковые номера сочетаний; по оси абсцисс — время в с; вертикальными штриховыми линиями А и Б и стрелками указаны соответственно момент светового раздражения и момент укола. Калибровка- 500 (верхний луч) и 250 (нижний луч) мкВ, 50 мс. Графики и осциллограммы представлены в едином временном масштабе.
процессов обучения, следует признать, что сами по себе изменения реакций нейронов при выработке временных связей в отсутствие дополнительных контрольных процедур еще не являются доказательством причастности тех или иных структур мозга к процессам обучения.
Как правило ищут корреляцию между изменением реакций нейронов и поведенческими сдвигами. Однако для заключения об организации системы, обеспечивающей условнорефлекторный процесс, надо по крайней мере знать: любые или вполне определенные изменения реакций нейронов соответствуют новой сигнальной
значимости сенсорных стимулов и в каких образованиях мозга эти изменения проявляются наиболее отчетливо. Для того чтобы установить связь нейронных перестроек именно с процессами «сцепления» условного и безусловного стимулов, необходимо исключить возможные другие причины. К таковым могут относиться: 1) возможность изменения эффективной интенсивности сенсорного стимула; 2) влияние на биоэлектрическую активность мышечных артефактов при позных перестройках и выполнении движений и 3) увеличение общей неспецифической возбудимости, так называемой тонической составляющей ассоциативного процесса (по Б. И. Котляру, 1977).
При регистрации реакций нейронов разных образований мозга во время формирования условного рефлекса возникает вопрос об их первичности. Конечные изменения на уровне одного нейрона могут быть одинаковыми независимо от первичности или вторичности их характера. Поиск наиболее существенных для обучения структур мозга заключается в определении локализации тех нейронных популяций, где изменения происходят в первую очередь. Критерием вовлеченности должен являться предшествующий характер реакций нейронов относительно поведенческой реакции, из чего логически вытекает необходимость существования «критических синапсов», в которых прежде всего происходят изменения, обусловливающие распространение импульсации по новому пути. Причем наряду с сигналъно-детерминиро-ванными реакциями описан класс сдвигов импульсной активности, которые также возникают по механизму воспроизведения следов ранее выработанных связей, но без непосредственного участия внешних сигналов — центрально-детерминированные реакции (М. Я. Рабинович, 1975). Именно эти реакции могут послужить удобной моделью для исследования механизмов произвольной двигательной деятельности.
Структуры мозга, в которых обнаруживаются первичные сдвиги нейрональной активности, согласно Дж. Олд-су, представлены в разных отделах мозга. Их можно разделить на неспецифические и специфические. К первой группе отнесены те образования, в которых нейроны
реагируют сходным образом на условные и любые сходные стимулы той же модальности, но отличающиеся от первых по своим физическим параметрам.
В специфических отделах нейроны не проявляют черт генерализации в реагировании и не имеют сходных реакций на новые стимулы. В качестве наиболее значимых для обучения отделов мозга отмечены фронтальная и сенсомоторная кора, а также гиппокамп, где обнаружены изменения импульсной активности пирамид слоя СА3 уже при первых сочетаниях условного и безусловного стимулов и разряд нейронов предшествовал двигательной реакции.
Группа американских ученых под руководством Ч. Вуди (1982) исследовала внутриклеточно реакции нейронов сенсомоторной коры кошек при выработке мигательного условного рефлекса на звук. Безусловным стимулом служило механическое постукивание по переносице. Реакции нейронов сенсомоторной коры предшествовали появлению первых мигательных движений. В результате обучения наблюдалось, с одной стороны, избирательное снижение порогов миографических реакций круговой мышцы глаза, осуществляющей мигание в ответ на поверхностную или внутрикорковую микростимуляцию сенсомоторной зоны, а с другой — избирательное усиление реакции и увеличение числа реагирующих нейронов, проецирующихся к соответствующей мышце. Было установлено, что пороги генерации спайков на внутриклеточно приложенный импульс тока достоверно ниже у тех нейронов, которые проецируются к мышцам, осуществляющим реакцию мигания, по сравнению с проекционными нейронами других лицевых мышц. Условные реакции нейронов на звуковой сигнал появляются с коротким латентным периодом, не превышающим латентный период реакций клеток первичной слуховой коры, и могут сохраняться в течение длительного времени — 14 недель.
Указанные функциональные сдвиги нейронов сенсомоторной коры были результатом только выработки условного рефлекса. Функциональное выключение сенсомоторной области аппликацией хлорида калия вызывает утрату реакций этих нейронов и соответственно
исчезновение самого акта мигания, хотя безусловная мигательная реакция не страдает. Результаты этих экспериментов свидетельствуют о первичности пластических изменений в нейронах коркового представительства двигательной условной реакции.
В пользу локализации пластических изменений в сенсомоторной коре свидетельствуют также результаты анализа нейронных механизмов условной реакции постановки конечности на опору у кошек (Б. И. Кот-ляр и др., 1983). Под этой реакцией понимают последовательность движений, вызванных прикосновением предмета к дорзальной поверхности лапы, обеспечивающих постановку лапы на поверхность этого предмета. Сигнальная условная реакция, вызванная тактильной стимуляцией вентральной поверхности передней конечности, имела то же двигательное проявление, что и исходная реакция, развивающаяся при раздражении дорзальной поверхности лапы (безусловная) у необученных животных.
В результате обучения возбудительный ответ нейронов в корковом представительстве бицепса плеча мышцы, выполняющей основную роль в сгибании локтевого сустава, на условный раздражитель был в 2—2,5 раза больше, чем ответ на тот же раздражитель у контрольных животных. Этот эффект не сопровождался изменением сенсорного притока в сенсомоторную кору.
Таким образом, реорганизация сенсомоторных связей, достигаемая путем обучения, состоит в изменении эффективности связей между нейронами сенсомоторной коры: 1) получающими сенсорно обусловленное эфферентное возбуждение и 2) контролирующими сокращениями определенных групп мышц.
Внутрикорковые системные механизмы временных связей. Технические условия ограничивают возможности одновременной регистрации активности большого числа нейронов. Чаще всего используют регистрацию активности одного, реже — двух—трех нейронов при формировании временных связей. Вместе с этим важно при анализе характера импульсации такого нейрона исходить из предпосылки его обязательного участия в составе определенной нервной сети.
У. Г. Гасанов (1981) полагает, что методическим приемом для выявления такой элементарной микросистемы является одновременная регистрация нескольких клеток с последующей выборкой отдельных импульсных потоков и статистический анализ перекрестных интервалов между импульсами двух и трех выделенных рядов.
Он выделяет следующие особенности взаимосвязной деятельности нейронов при обучении. 1. Деятельность микросети качественно отличается от индивидуальной активности входящих в его состав нейронов. 2. Системная деятельность корковых нейронов при разномодаль-ных длительных раздражениях, а также на разных стадиях выработки условного рефлекса и при угашении его основывается на разных формах взаимоотношений между нейронами. 3. Микросистема основывается на дифференцированном участии в ней различного класса нейронов, по-разному меняющих свои сетевые свойства при обучении.
Предполагается, что в основе изменений реакций нейрона при обучении лежат механизмы неспецифического характера. Локальные же микросети нейронов включаются в более обширные многоуровневые системы общецеребрального характера. Для выявления собственно корковых процессов формирования временных связей необходима либо внутриклеточная регистрация, которая по своей сути не может быть длительной, либо использование строго локальных воздействий на исследуемый нейрон, например с помощью микроэлектрофореза различных биологически активных соединений. Такая форма эксперимента предъявляет особые требования к самой процедуре условнорефлекторного обучения, состоящей в том, что и формирование, и угашение временной связи осуществляется на протяжении одного и того же опытного сеанса при параллельной регистрации импульсной активности одного и того же нейрона.
Мигательный условный рефлекс отвечает таким требованиям. В. С. Сабанов (1985) основное внимание обратил на самые ранние этапы формирования временной связи, когда внешних проявлений условной двигательной реакции еще не наблюдалось. При сочетании ритмического звукового и механического (задув воздуха на
роговицу глаза) раздражений наблюдалось два типа перестроек нейрональной активности: активация нейрона на условный сигнал, к которому данный нейрон был не реактивен, и тормозной: на условный сигнал появляется тормозная реакция, которая до сочетаний регистрировалась лишь в ответ на предъявление безусловного сигнала (рис. 56). Эта тормозная реакция вырабатывается быстро и угашается при изолированном действии условного сигнала. Характерно, что клетки с активирующим типом реакции были исходно полисенсорные, то есть реагировали на условный и безусловный сигналы, а клетки с тормозным типом реагирования на условный сигнал были исходно моносенсорными и становились полисенсорными лишь в процессе обучения. Исходно бисенсорные клетки с активирующими условными реакциями отражают результат ассоциативного процесса, сложившегося в результате сочетаний конвергирующих стимулов на докорковых уровнях интеграции, — статическая проецируемая конвергенция, тогда как тормозные условные реакции нейронов обусловлены внутрикорко-

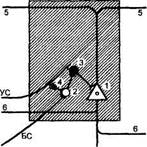
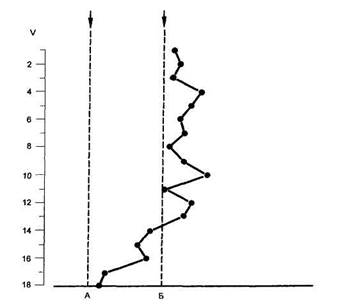
Рис. 57 Схема
связей нейронов для
объяснения происхождения
тормозных условных
клеточных реакций
1 — пирамидная клетка глубоких слоев коры; 2 — звездчатая клетка; 3 и 4 — тормозный интернейрон; 5 — ветви апикальных денд-ритов в поверхностном слое коры; 6 — коллатерали аксонов; УС — путь проведения условного стимула от соседнего нейронного модуля; БС — путь проведения безусловного стимула из таламуса.
выми механизмами конвергенции и интеграции — локальная динамическая конвергенция. Вопрос о месте формирования различного рода условных клеточных реакций имеет принципиальное значение.
Рассмотрим схему на рис. 57, из которой видно, что талами-ческий эфферент, передающий безусловную стимуляцию (БС), через звездчатую клетку, вступая в кору (2), приводит к активации тормозного интернейрона (3), непосредственно контактирующего с исследуемой крупной пирамидой (1). По мере выработки временной связи данный нейрон начинает реагировать аналогичной тормозной условной реакцией и на

