 2015-07-14
2015-07-14 1817
1817Дифференцировочное торможение развивается при неподкреплении раздражителей, близких к подкрепляемому сигналу. Этот вид торможения лежит в основе раз-
личения близких по своим физическим свойствам раздражений.
Представим себе следующую экспериментальную ситуацию. У собаки вырабатывается условный рефлекс на звуковой тон «ля». Все близкие к нему тоны при первом своем применении будут вызывать аналогичную реакцию. Это явление было названо И. П. Павловым генерализацией, то есть обобщением условных рефлексов. Однако, постоянно подкрепляя тон «ля» и не сопровождая действием безусловного раздражителя другие тоны, можно добиться их полного различения. Оно будет проявляться в наличии условной реакции только на тон «ля» и ее отсутствии на действие прочих тонов. С помощью дифференцировочного торможения организм из массы сходных раздражителей будет реагировать только на один подкрепляемый, то есть биологически для него важный, а на другие сходные раздражители условная реакция будет выражена слабее или отсутствовать полностью. Таким образом достигается после первичной генерализации условных рефлексов их последующая специализация.
При первом своем проявлении условный рефлекс может быть и не генерализован. Лишь по мере своего укрепления рефлекс приобретает генерализованный характер. Биологическое значение генерализации видно из следующего примера. Допустим, животному удалось избежать когтей хищника. Однако оборонительная реакция, совпадая по времени с голосом хищника, стала носить условнорефлекторный характер на звук голоса нападающего. Генерализация, или обобщение всех вариаций голоса хищника, которые возникают в естественной среде, обеспечивает выполнение оборонительной реакции в различных ситуациях, что является биологически выгодным для сохранения жизни организма. Специализация в этих условиях оказала бы пагубное влияние на животное, сильно ограничивая возможности его самозащиты. То же справедливо и для других видов поведения (пищевого, полового и пр.). Следовательно, свойство генерализации условных рефлексов — неизбежный атрибут поведенческих адаптации животных в естественной среде обитания.
Учитывая то обстоятельство, что изменчивость окружающей организм среды происходит по вероятностному закону, колебания тех или иных биологически значимых признаков предвидеть с высокой вероятностью невозможно, становится биологически оправданной значительная сенсорная генерализация условных рефлексов как стадии активного поиска жизненно важных объектов (Р. А. Павлыгина, 1983; П. В. Симонов, 1987). Р. А. Пав-лыгина считает, что в стадии генерализации условных рефлексов обнаруживается механизм доминанты, одной из характерных черт которой является способность доминирующей рефлекторной системы диффузно отзываться на самый широкий репертуар внешних стимулов, в том числе и впервые встречающихся. Однако известно, что постепенно, в процессе повторного осуществления данного рефлекторного акта, диффузная отзывчивость сменяется избирательным реагированием только на те раздражения, которые создали данную доминанту. Наступает стадия специализации доминанты, которая достигается благодаря механизмам дифференцировочного торможения. Таким образом, в создании доминанты как вектора целенаправленного поведения участвуют как процессы безусловного (сопряженного) торможения, так и процессы условного торможения.
Отметим следующие основные свойства дифференцировочного торможения: 1) чем ближе дифференцируемые раздражители, тем труднее на один из них (неподкреп-ляемый) выработать дифференцировочное торможение; 2) степень торможения определяется силой возбуждения, развиваемого положительным (подкрепляемым) условным рефлексом. Как и в случае угасательного торможения, дифференцировочное торможение легче вырабатывается при пищевом подкреплении, чем при оборонительном, легче у накормленных животных, чем у голодных; 3) выработка этого торможения происходит волнообразно; 4) дифференцировочное торможение тренируемо, что лежит в основе тонкого распознавания "сенсорных факторов среды.
В самостоятельный вид условного торможения И. П. Павлов выделил условный тормоз, который образуется при неподкреплении комбинации из положи-
тельного условного раздражителя и индифферентного. Например, у собаки образован пищевой условный рефлекс на звук. Если к этому сигналу присоединить свет от лампочки и их совместное действие не подкреплять пищей, то после нескольких применений этой комбинации она перестанет вызывать пищевую реакцию. Хотя изолированное применение звонка по-прежнему будет вызывать обильное слюноотделение.
По-существу — это вариант дифференцировочного торможения. Этот вид торможения часто встречается. Например, при хозяйке домашняя кошка ведет себя «благоразумно», а в отсутствие ее забирается на стол и поедает пищевые продукты. В этом случае вид стола — условный пищевой сигнал, сочетание вида стола и хозяйки никогда не сопровождалось пищей и сделалось условным тормозом. Присутствие хозяйки (условного тормоза) подавляет пищедобывательную реакцию кошки на вид стола. Выработанные в течение жизни условные тормоза, как и дифференцировочное торможение, уточняют условные рефлексы, делают поведение более приспособленным к конкретной окружающей ситуации.
Основные свойства условного тормоза следующие.
1. Он легче вырабатывается, если к слабому (световому, например) положительному раздражителю присоединяется сильный дополнительный, например звонок.
2. Роль прибавочного раздражителя может играть след от примененного раздражения достаточно большой силы. Так, след от автомобильного гудка даже спустя 20 с может служить в качестве условного тормоза к мет-рономному сигналу условного пищевого рефлекса.
3. Если прибавочный раздражитель имеет недостаточную силу, то он может превратиться в условный раздражитель второго порядка (см. гл. 10), то есть выступить в качестве положительного сигнала основного условного рефлекса.
4. Если прибавочный раздражитель приобрел свойства условного тормоза, то, будучи присоединенным к любому другому положительному сигналу, он затормозит соответствующий этому сигналу условный рефлекс.
5. Прибавочный раздражитель в первый момент своего применения в комбинации с положительным сигналом
9 Зак S 64
вызывает ориентировочный рефлекс и индукционное торможение условной реакции, затем превращается в индифферентный раздражитель (гаснущий тормоз), и, наконец, на месте безусловного торможения развивается условный тормоз.
Торможение запаздывания. При выработке такой разновидности условного торможения подкрепление соответствующим безусловным рефлексом не отменяется (как в предыдущих видах торможения), а значительно отодвигается от начала действия условного раздражителя. Подкрепляется лишь последний период действия сигнала, а предшествующий ему значительный период его действия лишается подкрепления. Поэтому первый период действия условного сигнала, который сопровождается торможением запаздывания, называется недеятельной фазой запаздывающего условного рефлекса. По истечении его торможение прекращается и сменяется возбуждением — так называемая деятельная фаза рефлекса.
Строго говоря, в данном случае действует не один раздражитель, а два в комплексе, и второй компонент — это время. Именно время определяет момент прекращения запаздывательного торможения и наступление деятельной фазы рефлекса. Допустим, у собаки выработан запаздывающий условный рефлекс на свет. Экспериментатор зажигает электрическую лампу, которая до момента подкрепления горит 3 мин. Условнорефлектор-ная реакция, возникавшая по световому сигналу, не подкрепляется и поэтому быстро гаснет, так как развивается угасательное торможение. Но за некоторое время до подкрепления условная реакция проявляется четко, она подкрепляется. Таким образом, условная реакция как бы приурочивается к моменту действия безусловного раздражителя. Начальный период действия раздражителя выступает в качестве сигнала отрицательной реакции, а более длительный промежуток времени, близкий ко всему периоду запаздывания, служит положительным сигналом. В случае пищевых условных рефлексов отставление подкрепления от начала действия условного сигнала может достигать 2—3 мин, а при электрооборонительных двигательных рефлексах этот интервал значительно короче — 30-60 с.
Глава 6. Закономерности условно рефлекторной деятельности 259
Адаптивное значение торможения запаздывания состоит в тонком анализе времени отставления раздражителя; условный рефлекс более или менее точно приурочивается ко времени действия сигнализируемого безусловного рефлекса.
Отметим основные свойства данного вида условного торможения.
1. Чем сильнее условный раздражитель, тем труднее выработать торможение запаздывания.
2. Чем больше сила подкрепляющего рефлекса, тем труднее выработать запаздывание условного рефлекса.
3. Чем медленнее удлиняется от опыта к опыту изолированное действие условного раздражителя, тем легче вырабатывается запаздывание. Если подкрепление сразу отодвинуть от начала действия положительного сигнала на 2-3 мин, то запаздывание выработать чрезвычайно трудно.
4. Значительное упрочение совпадающих или корот-коотставленных условных рефлексов препятствует выработке торможения запаздывания.
Хотя торможение запаздывания и отличается от трех других видов условного торможения благодаря участию временного фактора, в основе его развития также лежит неподкрепление определенного периода действия условного сигнала.
Взаимодействие разных видов торможения. Взаимодействие условного и безусловного торможения может происходить чаще всего в форме феномена растормажи-вания, о котором уже упоминалось. Суть его состоит в том, что безусловное индукционное торможение, возникающее при ориентировочном рефлексе, временно ослабляет или полностью уничтожает эффект условного выработанного торможения (угасательного, дифферен-цировочного, условного тормоза и торможения запаздывания). Например, животное обучилось выполнять защитное поведение только на определенные звуки своего врага и не реагировать на все остальные. Но выйдя на очередную охоту в привычной для себя обстановке, животное встретилось с неожиданным новым запаховым раздражителем, который естественно вызвал ориентировочный рефлекс. Одновременно с этим настороженность
животного сильно возросла, ибо теперь уже ранее угашенные звуковые раздражители начинали вызывать защитную реакцию (затаивание и убегание). Из такого примера ясно, что феномен растормаживания имеет важное биологическое значение, обеспечивая срочную перестройку привычного поведенческого стереотипа к новой жизненной ситуации. Конечно, эффект растормаживания очень динамичен, он зависит от многих факторов: силы и прочности выработанного торможения, силы и продолжительности ориентировочного рефлекса, возраста и типа нервной системы животного.
Другое направление взаимодействия разных видов торможения состоит в их суммации. Например, угаше-ние условного рефлекса может ускоряться, если кроме неподкрепления применить умеренной силы посторонний раздражитель. Происходит суммация условного уга-сательного и безусловного индукционного торможения.
Не меньше примеров еуммации разных видов условного торможения; скажем, одновременная тренировка угасательного и запаздывательного торможения взаимно усиливают друг друга. Или в другом случае один из видов условного торможения (например, дифференциро-вочное) формируется быстрее, если ему предшествовала процедура с тренировкой угасательного торможения. Биологическое значение эффекта суммации торможения понятно, если иметь в виду, что тренировка торможения — одно из основных условий формирования и воспитания различной сложности поведенческих навыков.
Сами факты взаимодействия разных видов торможения являются достаточным основанием, чтобы предполагать их близкую физиологическую природу.
§ 32. ЗАКЛЮЧЕНИЕ
Наряду с относительно постоянными формами реагирования на стабильные внешние и внутренние воздействия (безусловные рефлексы) существует другой тип реакций, позволяющих организму адекватно реагировать на все изменения окружающей среды (условные рефлексы). Условные рефлексы образуются при совпадении во времени и пространстве какого-либо индифферентного
(условного) раздражителя и раздражителя, вызывающего безусловный рефлекс. Главный смысл условного рефлекса в его сигнальности (предупреждающем характере) и адаптивности (приспособительном характере). Условные рефлексы разделяются по признаку особенностей безусловного подкрепления, условного сигнала или по временному соотношению обоих сигналов.
И. П. Павлов разработал классификацию феноменов торможения условных рефлексов. К внешнему (безусловному) торможению он отнес гаснущий и постоянный тормозы, а также запредельное торможение. В первых двух случаях условные рефлексы тормозятся при действии ориентировочного или болевого раздражений. К внутреннему (условному) торможению, которое возникает вследствие неподкрепления условного сигнала безусловным рефлексом, относят угасательное торможение, дифференцировочное, запаздывания, условный тормоз.
По-видимому, сходство физиологической природы разных типов и видов торможения условных рефлексов не исключает различия в их нейрофизиологических механизмах.
262
Глава 7. МЕХАНИЗМЫ ФОРМИРОВАНИЯ УСЛОВНЫХ РЕФЛЕКСОВ
§ 33. КОНВЕРГЕНТНАЯ ТЕОРИЯ ФОРМИРОВАНИЯ ВРЕМЕННЫХ СВЯЗЕЙ
Г азработке основных законов центральной нервной координации физиология обязана исследованиям Ч. Шеррингтона, сформулировавшего фундаментальный закон в работе нервной системы — «принцип общего пути». Если афферентный нейрон является индивидуальным или частным путем, то эфферентный нейрон является общим путем для импульсов, возникающих в любом из многочисленных источников рецепции. Подобная координация складывается на основе морфологического соотношения афферентных и эфферентных путей, где первые всегда значительно преобладают над вторыми. Так формируется и реализуется механизм конвергенции — схождения множества афферентных импульсов в единственный анатомически ограниченный эфферентный канал. Частные пути, конвергируя между собой, соединяются в промежуточные пути и окончательно конвергенция завершается общим конечным путем в виде воронки. Каждый этап, на котором два или более афферентных нейрона конвергируют на третьем нейроне, который по отношению к ним является эфферентным, предполагает возможность влияния или столкновения их влияний.
Идею о принципе конвергенции творчески развил А. А. Ухтомский (1954), распространив его на всю центральную нервную систему, на все ее этажи. Он полагал, что в результате конвергенции наступает интерференция стимулов, которая реализуется в определенный нервный акт. Отсюда становится понятным богатое развитие синапсов и концевых ветвлений с тенденцией уве-
263
личить поверхность соприкосновения нервной клетки с окончаниями приходящих аксонов.
Автор принципа доминанты выдвинул новые положения об интегрирующей деятельности нервной системы. Принцип статической конвергенции путей по отношению к исполнительному пути при дальнейшей разработке расширяется в принцип подвижной и физиологической конвергенции нервных импульсов по отношению к области преобладающего текущего действия (принцип доминанты).
А. А. Ухтомский находил тесную связь между учением Шеррингтона об общих путях и теорией И. П. Павлова об условных рефлексах. В статье «Об условно-отраженном действии» он писал: «Если воронка Шеррингтона имела в виду постоянные рефлекторные дуги, закрепившие функциональную и морфологическую связи между собой наследственно и филогенетически, то И. П. Павлов улавливал самое закладывание и новообразование связей в этой воронке, привлечение все новых рецептивных сфер к конвергенции относительно эфферентных (исполнительных) приборов».1
Конвергенция гетеросенсорных стимулов была положена в основу понимания механизмов образования условных рефлексов. Так, Дж. Экклс (1953) предположил существование нелокализованного «центра конвергенции», где сходятся условный и безусловный раздражители. Встреча их в центре конвергенции, взаимодействие и интеграция завершаются формированием особой пространственно-временной структуры разрядов, которая затем передается в нейронную сеть. Многократные сочетания включают механизмы потенциации путей условного сигнала, который приобретает способность запускать условнорефлекторный акт. Длительность хранения этого эффекта обеспечивается механизмами реверберации возбуждения в нервных сетях. А. Фессар (1962) показал, что конвергенция, то есть сближение в пространстве мозга разномодальных сигнализаций, является важнейшим условием формирования временной связи на уровне отдельных нейронов. В качестве двух других условий он
1 Ухтомский А. А. Доминанта. Л., 1966. С. 175.
выдвинул взаимодействие этих сигнализации на нейронах и использование следовых процессов. Взаимодействие между конвергирующими сигналами связано с длительным удерживанием следов активности. В этих процессах важную роль играют механизмы тонической и пост-тетанической потенциации.
Таким образом, конвергенция и потенциация образуют некую общую цепь динамических процессов, связывающих интеграцию в пространстве (обусловленную конвергенцией) с интеграцией во времени (обусловленную постепенным нарастанием облегчения на окончаниях аксонов) (А. Фессар, 1964).
Первичная конвергенция осуществляется преимущественно на клетках ретикулярных образований, та-ламуса и ствола мозга и передается на корковые нейроны в интегративной форме («проецируемая мульти-валентность»). Корковые структуры регулируют конвергентные свойства ретикулярных структур, что обеспечивает специализацию условного рефлекса (рис. 48). Эти концепции не могли объяснить свойственные условному рефлексу устойчивость и приспособительную пластичность, ибо не придали должного значения фактору биологического подкрепления. Именно это было учтено П. К. Анохиным (1968), который считал, что решающим фактором в механизмах образования условных рефлексов выступает биологическое значение подкрепляющего рефлекса.
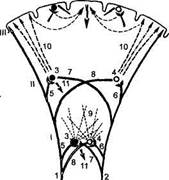
Рис. 48
Схема образования условного рефлекса (по А. Гасто, 1958)
I — ретикулярная формация нижней части ствола мозга; // — таламическая ретикулярная формация; III — кора мозга; 1 — афферентный путь безусловного раздражителя; 2 — афферентный путь условного раздражителя; 3 и 4 — ретикулярные нейроны; 5 и 6 — коллатеральные волокна афферентных путей, вызывающие возбуждение ретикулярных нейронов; 7 и 8 — коллатеральные волоква афферентных путей, не вызывающие возбуждения нейронов вследствие непроходимости синапсов; 9 — восходящие пути от ретикулярной формации, активирующие кору диффузно; 10 — восходящие пути от ретикулярной формации, активирующие кору локально; И — безуслов-норефлекторная реакция. Временная связь образуется между афферентными влияниями условного и безусловного раздражителей.
Конвергирующие на нейронах сенсорные и биологические модальности определяют организацию условного рефлекса — его сигнальные свойства и приспособительное значение. Стимулы различной сенсорной и биологической модальности воздействуют при сочетаниях на синаптические входы нейронов с гетерохимической организацией этих входов. Свойства подкрепляющего фактора определяются специфическими химическими реакциями, которые он вызывает, и соответствующими молекулярными преобразованиями в пределах мембраны и цитоплазмы нейрона. Здесь и происходит «сцепление» условной и безусловной стимуляции с последующим выходом сложившейся интеграции на аксон в виде клеточного разряда.
Само понятие «сцепление» следов условного и безусловного стимулов предполагает наличие специфических ультраструктурных и молекулярных преобразований в цитоплазме нейрона. Гипотетическая схема, предложенная М. Я. Рабиновичем (1975), приведена на рис. 49. Сочетание стимулов двух сенсорных модальностей (бг и б2) приводит к образованию кратковременной связи типа сенсо-сенсорной ассоциации. Сочетание же каждого из сенсорных стимулов с биологически значимым подкреплением (а) эмоционально-мотивационного характера приводит к выработке устойчивой связи, обладающей свойствами условного рефлекса.
Признавая, что наиболее существенные процессы образования временных связей реализуются на молекулярном
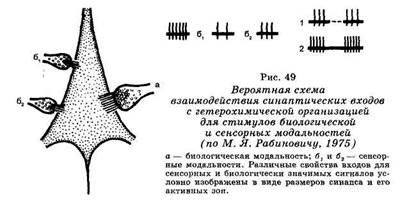
уровне нейрона, следует иметь в виду, что системные механизмы временных связей всецело опосредованы сложными нейрофизиологическими преобразованиями. Выделяют три типа проявлений мультисенсорной конвергенции (Л. Л. Воронин, 1974). Первым типом является «специфический ассоциативный», при котором клетки ствола мозга, таламуса или коры реагируют специфическим ответом на стимулы разных сенсорных модальностей. Вторым типом является «специфический проекционный», при котором клетка реагирует специфической реакцией на адекватный для данной системы раздражитель и неспецифической — на стимулы других модальностей. Наконец, третий тип именуется неспецифическим и характеризуется неспецифической ответной реакцией нейронов на разномодальные стимулы. М. Я. Рабинович (1975) разработал гипотетическую схему, в которой синаптические входы для условного и безусловного стимулов конвергируют на пресинаптичес-
Рис. 50
Гипотетическая схема передачи условной связи, сложившейся
в процессе сочетаний, с «входного нейрона» (А.) на выходной нейрон
(Б) по одному общему пути (по М. Я. Рабиновичу, 1975)
Клетка круглой формы — вставочный нейрон возбуждающего действия; на осциллограммах — различные формы условной активности, которые возможно регистрировать с «выходного» нейрона: / — обычная реакция на условный и безусловный стимулы (УС и ВС), обозначенные длинной и короткой линией под осциллограммой; 2 — воспроизведение следовых разрядов на БС при его пропуске в сочетании; 3 — воспроизведение следовых разрядов на УС и БС при пропуске всего сочетания (обозначено пунктиром) в условиях реакции на «время».
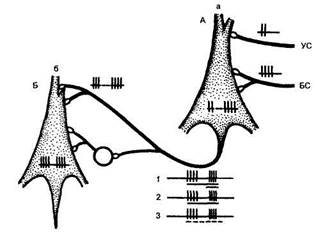
кой («входной») клетке в цепи нейронов. Сложившаяся ассоциация («интеграция») передается на постсиналти-ческий нейрон («выходной»), но по одному общему пути и для сигнального, и для подкрепляющего раздражителей (рис. 50).
При рассмотрении разномодальной конвергенции необходимо учитывать чрезвычайно важное свойство нейронов и нервных сетей, а именно способность к организации пластического характера конвергенции. В этой связи следует разграничивать конвергенцию анатомическую, или статическую, и конвергенцию функциональную, или динамическую. Особенности организации нервных сетей головного мозга обеспечивают связь по принципу «от каждого ко всем». Следовательно, морфологические предпосылки могут создать такие условия, когда на одном нейроне будут конвергировать самые разнообразные влияния. В процессе эволюции и в динамике постнатального онтогенеза в головном мозге закрепляется олигосинаптическая сеть, обеспечивающая четкий адресат проведения информации наряду с системой по-лисинаптических сетей, связи между компонентами которой могут носить функциональный временный характер. Это обстоятельство, то есть высокая вариативность свойств полисенсорных нейронов, и обусловливает многообразие их реакций на разномодальную стимуляцию.
§ 34. КЛЕТОЧНЫЕ АНАЛОГИ УСЛОВНОГО РЕФЛЕКСА
Выделяют две основные гипотезы обучения — синоптическую, предполагающую, что механизмом формирования условных рефлексов является изменение эффективности синапсов, и мембранную, утверждающую, что основным механизмом условного рефлекса является изменение свойств возбудимой постсинаптической мембраны. Как мы увидим далее, такое деление весьма условно, однако представляется удобным для первого знакомства с проблемой.
Нервные механизмы обучения обеспечиваются благодаря двум основным свойствам нервной системы — реактивности и пластичности. Если реактивность -- это
способность отвечать на раздражения, то пластичность — это способность изменять реактивность под влиянием последовательных раздражений. В свою очередь реактивность определяется возбудимостью постси-наптической мембраны и эффективностью соответствующих синапсов. Возбудимость нейрона измеряется силой тока, необходимой для генерации потенциала действия, а синаптическая эффективность — амплитудой моно-синаптического постсинаптического потенциала.
Уже давно высказывалось предположение о том, что в основе обучения лежит изменение эффективности межклеточных контактов или проторение новых контактов. На рис. 51 приведены основные схемы. Простейшим является синапс Дж. Экклса (А), повышающий свою эффективность после собственной активации (фасилита-ция). Такие синапсы могут служить для объяснения феноменов посттетанической потенциации. Дополнением к ним являются синапсы с постепенным снижением эффективности при повторной активации (В). Они могут обеспечивать эффекты габитуации и дегабитуации. Как мы уже отмечали, при выработке условных рефлексов необходимо временное совпадение пресинаптической активации с активацией подкрепляющих систем, которые могут оказывать также пресинаптическое действие (В). Д. О. Хебб (1959) главным условием синаптической модификации считал совпадение пресинаптической актив-
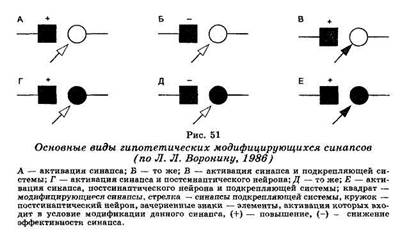
ности с разрядом постсинаптического нейрона (Г). Затем к этому была добавлена модель Альбуса, связанная не с увеличением эффективности синапса, а с ее снижением (Д). Наконец, в качестве синтетической схемы (Е) рассматриваются случаи изменений синаптической эффективности с участием трех факторов — сенсорного притока, активации подкрепляющей системы и достижения результата.
Вопрос о характере преобразований нейрональной активности в динамике выработки условного рефлекса начал исследоваться в связи с внедрением в практику физиологического эксперимента тонких методов микроэлектродного анализа и последующей компьютерной обработки импульсных реакций нейронов. Попытки использовать внутриклеточную регистрацию постсинаптических электрических потенциалов привели к необходимости проводить исследования либо на относительно простых нервных сетях, либо на центральных нейронах высших млекопитающих с максимальным упрощением требуемых для анализа межнейронных отношений. Такие упрощенные модели принято рассматривать в качестве клеточных аналогов условного рефлекса.
Группой венгерских физиологов (О. Фехер, А. Бара-ни, 1981), разработана модель ассоциативного обучения, при котором сигнал меняет свое действие под влиянием последующего безусловного эффекта.
Рассмотрим процедуру обусловливания при «условном» раздражении вентролатерального ядра таламуса (VL) и антидромной стимуляции пирамидного тракта («безусловный стимул»). Сначала, как показано на рис. 52, электрическая сшмуляция VL с частотой 1/с сопровождается только яодпороговыми ПСП. Последующее антидромное раздражение пирамидного тракта вызывает один потенциал действия. По мере сочетаний раздражения таламуса с антидромной стимуляцией амплитуда и/или длительность ТПСП увеличиваются и остаются таковыми в течение нескольких минут. Амплитуда ВПСП изменяется также, но к 50-му сочетанию ВПСП достигает порога генерации и «условный» сигнал начинает вызывать 1-2 потенциала действия. Эффект антидромного раздражения при этом не меняется. К 100-му сочетанию
амплитуда ВПСП еще больше возрастает, и число потенциалов действия достигает трех. Если проводить угаше-ние, то есть подавать одно «условное» раздражение, не сопровождая его антидромной стимуляцией, то амплитуда ВПСП и число генерируемых потенциалов действия уменьшается, и спустя 350 изолированных раздражений таламического ядра ответная «условная» реакция исчезает. Внутриклеточная поляризация исследуемого нейрона, приводившая к генерации потенциалов действия, усиливала эффект раздражения вентролатерального та-ламуса. Однако ни многократное применение только «условного» стимула, ни псевдообусловливание (процедура применения условного и безусловного раздражителей в случайном порядке) не привели к генерации потенциалов действия исследуемого нейрона. Уровень мембранного потенциала и сопротивление мембраны при выработке временной связи менялись по-разному в различных клетках. Это позволило думать об определяющей роли при выработке временной связи избирательных синаптических сдвигов.
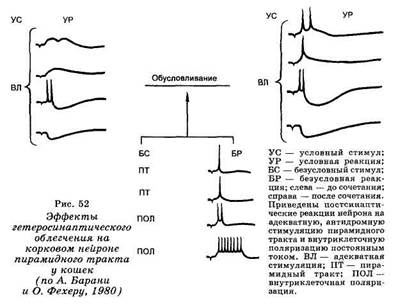
Было показано, что положительные и дифференци-ровочные связи с исследуемым нейроном опосредуются разными синапсами независимо друг от друга. Е. Н. Соколов (1981) справедливо полагает, что элементарным локусом пластического изменения является не нейрон, а синапс. Речь идет о синапсе Хебба, который селективно изменяется, если за синаптической посылкой следует спайковый разряд нейрона. Такое явление получило наименование гетеросинаптического облегчения.

