 2015-10-13
2015-10-13 3123
3123В настоящее время общепринятой является точка зрения, что колониальное гнездование у птиц возникло в процессе эволюции из одиночно-территориального. Однако, обращает на себя внимание то, что колониальный способ гнездования доминирует в наиболее древних отрядах — пингвинов, трубконосых, веслоногих, голенастых, ржанкообразных. В первых четырех колониальные виды aбсолютно доминируют, составляя в каждом от 85 до 100%. Это позволяет предположить, что у части птиц эволюция шла от колониального гнездования к одиночно-территориальному, то есть для некоторой части отрядов птиц первичным был колониальный способ гнездования. Как косвенное подтверждение данной гипотезы можно привести мнения многих авторов о том, что колониальное гнездование не дает особых преимуществ при размножении. Ecли сравнить птиц, гнездящихся облигатно, с птицами, которым присущ факультативный способ гнездования, то выясняется, что облигатный способ гнездования в основном характерен для древних видов.
Рассматривая характеристику облигатного типа гнездования, данную В.А.Зубакиным (1976) для отряда ржанкообразных.
Отсутствие охраняемой территории — охраняется только само гнездо,
криптичность яиц понижена,
дефекация птицы на край гнезда;
отсутствие выноса скорлупы из гнезда после вылупления птенцов,
образование птенцами табунков или затаивание на гнезде,
каннибализм,
транспорт эктопаразитов и возможность вспышек эпизоотии в результате скученности,- следует признать, что все вышеперечисленные признаки вполне можно трактовать как проявление примитивности, то есть облигатно-колониальный способ гнездования, возможно, является наиболее древней формой гнездования из ныне существующих.
При факультативном типе гнездования
- птицы имеют охраняемую гнездовую территорию, различную по величине у разных видов,
- способны успешно гнездиться отдельными парами,
- маскируют гнезда,
- дефекация в гнездо отсутствует,
- после вылупления птенцов скорлупа из гнезда удаляется
Т.е. одиночно-территориальное гнездование очень близко к факультативно-диффузному и сохраняет большинство черт, присущих данному способу гнездования, и, скорее всего, является следующим этапом в процессе эволюции.
В наиболее современном отряде воробьиных колониальность присуща только приблизительно 6% видов, при этом у некоторой части видов отряда воробьиных она, возможно, вторичного происхождения.
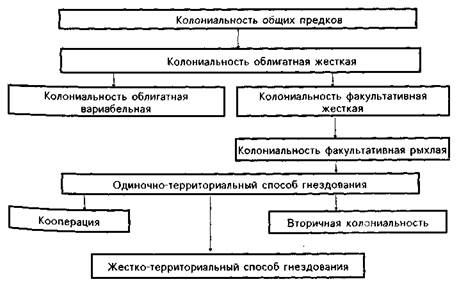
Конечно, эволюционный процесс проходил в разных группах с разной скоростью, остановками, параллелизмом, вариациями направлений, вероятно, на определенных этапах носил в разных отрядах противоположную направленность, но общий его ход, нам кажется, можно представить так:
Обследуя колонии голенастых на восточном Сиваше, наблюдали, как первыми заселяли массивы тростниковых зарослей большие белые и серые цапли, образуя рыхлые колони где гнезда находились друг от друга в 10—50 м. Следом прилетали рыжие цапли и поселялись на участке, уже занятом серыми большими белыми цаплями, колонии уплотнялись и увеличивались в размерах. Затем в колонии вселялись колпицы, кваквы, каравайки, следом желтые и малые белые цапли, при этом в тростниковых зарослях трехметровой высоты образовались плотные трехъярусные поселения, поливидовые по составу. Гнезда почти касались краями друг друга, то есть возникли очень плотные поселения, центры которых представляли структуры, близкие к колониям облигатного типа, с переходом на периферии к разреженному гнездованию. Если вспомнить, что у цапель отсутствует охраняемая территория у гнезда, а охраняется только само гнездо, дефекация при насиживании происходит на край гнезда, криптичность яиц понижена, скорлупа после вылупления не выносится, птенцы во время тревоги затаиваются на гнезде или пытаются; его оборонять, в более взрослом возрасте разбегаются, то выясняется, что в норме реакций у голенастых, при сохранении облигатного типа гнездования, плотность поселений может варьировать в зависимости от места гнездования и видового состава колоний. Эту форму колониальности мы считаем облигатно-вариабельной.
Таким образом, кроме общепризнанного пути эволюции колониальности у птиц — от одиночного гнездования к колониальному, — существовал еще один: от колониального к одиночно-территориальному. При этом в разных группах эволюция, возможно, на определенных этапах шла в противоположных направлениях. Наличие в древних отрядах птиц как облигатного способа гнездования, так и факультативного и даже одиночно-территориального говорит о том, что разветвление путей эволюции гнездования у птиц произошло на очень ранних стадиях развития.
Причины колониальности, выдвинутые Модестовым, — скопление пищевых ресурсов, дефицит мест гнездования и влияние хищников — можно рассматривать не как причины, а как факторы, оценивающие колониальность на жизнестойкость. Исследуя поливидовые колонии, нужно учитывать, что такие структуры представляют собой сумму репродуктивных стратегий составляющих их видов, где успешность размножения зависит как от видового состава, так и от общей численности особей в данных поселениях. Очевидно, что для птиц, гнездящихся колониально, существует оптимальная численность особей в поселении, при котором успешность размножения наибольшая.
Интересным представляется вопрос о предельной численности колонии т.е предельной численности вида в колонии. Предельная численность колонии для разных видов различна, но она существует, так как в однородных биотопах колонии не представляют собой сплошные поселения, а делятся на субколонии. При дефиците мест гнездования и большой численности вида в регионе этот вид может образовывать поселения, значительно превышающие оптимальную численность, где количество птиц достигает предельной величины: в этом случае происходит резкое снижение успешности размножения.
Интересно отметить, что эффект Дарлинга срабатывает до определенной величины поселения, за пределами которой синхронизации цикла размножения не происходит. Наличие механизма, регулирующего общую численность и численность отдельных видов в колонии, позволяет предположить, что колония — саморегулирующаяся система, стремящаяся к оптимальному состоянию. Механизмом саморегуляции, возможно, являются уровни голосовых и визуальных контактов внутри колонии. В пользу этого предположения говорит то, что в поселениях такие «молчаливые» птицы, как каравайки и бакланы, издают звуки, то есть в поселениях всегда существует звуковой фон.
По поводу гипотезы «колония как информационный центр» высказываются противоречивые мнения. На наш взгляд, представляется весьма сомнительным наличие системы передачи сложной информации у птиц с неразвитой голосовой коммуникацией. К этой группе относятся пингвины, трубконосые, веслоногие, голенастые, то есть наиболее древние группы с первичной колониальностью. Хотя простая информация передается: так, тревожный крик серебристой чайки вызывает массовый взлет хохлатых бакланов с колонии; у голенастых в поселениях звуковой фон складывается из специфических звуков, издаваемых разными видами цапель; в случае опасности птицы замолкают, после нескольких секунд тишины следует массовый взлет птиц с колонии — отсутствие комфортных звуков служит сигналом опасности. У ржанкообразных (на примере Майковых) уже проявляются некоторые признаки зачатков информационной системы. Наиболее вероятно функционирование колонии как информационного центра у воробьиных с высокой степенью вторичной колониальности. Примером такого центра может служить колония розовых скворцов. Поселение насчитывало около 4 тысяч пар и располагалось в каменной осыпи, в колонии птицы постоянно поддерживали друг с другом голосовой контакт, вылет за кормом и возвращение осуществлялись почти исключительно стаями от 20 до 150 особей, через определенные промежутки времени. На кормежку и с кормежки скворцы следовали узким коридором, четко сформированными стаями, место кормежки (очаг саранчи) находилось в 15—20 км от поселения. Отдаленность места сбора корма и пищевая специализация предполагают обязательную разведку мест концентрации пищевых объектов и передачу об этом информации, В данном случае в разведанный очаг саранчи летало до 60% птиц, еще около 30% особей приносили корм из очага размножения гусениц, остальные 10% собирали корм в окрестностях поселения. Кормежка происходила очень организованно, методом облавы, птицы во время нее постоянно перекликались, взлеты и посадки осуществлялись синхронно. Появление наземного хищника в колонии вызывало тревогу в локальном участке колонии, при этом голосовая коммуникация позволяла ориентироваться птицам не только в верхнем ярусе, но и в глубине осыпи. Из вышеизложенного видно, что значение колонии как информационного центра увеличивается по мере перехода птиц от первичной колониальности к вторичной.








