 2018-01-21
2018-01-21 13110
13110В ее состав входят: (1) синусный узел (который также называют синоатриальным или С-А узлом), где и происходит ритмическая генерация импульсов; (2) предсердные межузловые пучки, по которым импульсы проводятся от синусного узла к агриовентрикулярному узлу; (3) атриовентрикулярный узел, в котором происходит задержка проведения импульсов от предсердий к желудочкам; (4) атриовентрикулярный пучок, по которому импульсы проводятся к желудочкам; (5) левая и правая ножки А-В пучка, состоящие из волокон Пуркинье, благодаря которым импульсы достигают сократительного миокарда.
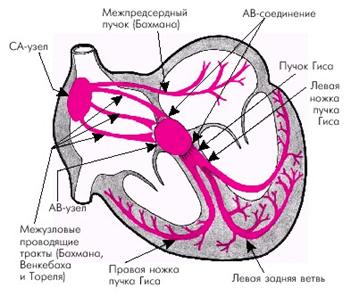
Проводящая система сердца образована двумя видами специализированных клеток.
Один вид клеток (Р-клетки) обладает автоматизмом, т.е. способностью спонтанно вырабатывать электрические импульсы. Частота возникновения ипульсов зависит от места расположения Р-клеток - чем ближе находятся Р-клетки к началу проводящей системы, тем чаще возникают в них импульсы и, наоборот, чем дальше находятся Р-клетки от начала проводящей системы, тем реже могут возникать в них электрические импульсы.
Второй вид (Т-клетки) обладает проводимостью, т.е. способностью проведения возникающих импульсов к сократительному миокарду.
Проводящая система сердца начинается синусовым узлом, который расположен в верхней части правого предсердия. Его длина 10-20 мм, ширина 3-5 мм. Именно в нем возникают импульсы, которые вызывают возбуждение и сокращение всего сердца. Нормальный автоматизм синусового узла составляет 50-80 импульсов в минуту. Синусовый узел является автоматическим центром I порядка.
Импульс, возникший в синусовом узле мгновенно распространяется по предсердиям, заставляя их сократиться. Но распространиться дальше и сразу же возбудить желудочки сердца эта волна не может, так как миокард предсердий и желудочков разделен фиброзной тканью, которая не пропускает электрические импульсы. И только в одном месте этой преграды не существует. Туда и устремляется волна возбуждения. Но именно в этом месте находится следующий узел проводящей системы, который называется атриовентрикулярным (длина около 5 мм, толщина - 2 мм). В нем происходит задержка волны возбуждения и фильтрация входящих импульсов.
Далее нижняя часть узла, утончаясь, переходит в пучок Гиса (длина 20 мм). В последующем пучок Гиса разделяется на две ножки - правую и левую. Правая ножка проходит по правой стороне межжелудочковой перегородки и разветвляясь ее волокна (волокна Пуркинье) пронзают миокард правого желудочка. Левая ножка проходит по левой половине межжелудочковой перегородки и делится на переднюю и заднюю ветви, которые снабжают волокнами Пуркинье миокард левого желудочка. После задержки в результате прохождения атриовентрикулярного узла волна возбуждения, распространяясь по ножкам пучка Гиса и волокнам Пуркинье, мгновенно охватывает всю толщу миокарда желудочков, вызывая их сокращение. Задержка импульса имеет огромное значение и не дает сократиться предсердиям и желудочкам одновременно - сперва сокращаются предсердия, и только вслед за этим - желудочки сердца.
В атриовентрикулярном узле, так же как и в синусовом узле, имеются два вида клеток - Р и Т. Атриовентрикулярный узел вместе с начальной частью пучка Гиса является автоматическим центром II порядка, который может самостоятельно вырабатывать импульсы с частотой 35-50 в минуту.
Конечная часть пучка Гиса, его ножки и волокна Пуркинье также обладают автоматизмом, однако могут вырабатывать импульсы лишь с частотой 15-35 в минуту и являются автоматическим центром III порядка.
Между автоматическими центрами I, II и III порядков возникают следующие взаимодействия. В норме импульс, возникающий в синусовом узле, распространяется на предсердия и желудочки, вызывая их сокращения. Проходя на своем пути автоматические центры II и III порядков импульс каждый раз вызывает разрядку этих центров. После этого в автоматических центрах II и III порядков снова начинается подготовка очередного импульса, которая каждый раз вновь прерывается после прохождения возбуждения из синусового узла. По сути дела, в норме автоматический центр I порядка подавляет активность автоматических узлов II и III порядков. И только в случае отказа синусового узла или нарушения проведения его импульсов на нижележащие отделы включается автоматический узел II порядка, а при его отказе - автоматический узел III порядка.
22.Потенциал действия в клетках рабочего миокарда.
Клетки проводящей системы сердца и, в частности, клетки пейсмекера, обладающие автоматией могут спонтанно деполяризоваться до критического уровня. В таких клетках за фазой реполяризации следует фаза медленной диастолическои деполяризации (МДД), которая приводит к снижению МП до порогового уровня и возникновению ПД. МДД — это местное, нераспространяющееся возбуждение, в отличие от ПД, который является распространяющимся возбуждением. Таким образом, пейсмекерные клетки отличаются от кардиомиоцитов: 1) низким уровнем МП — около 50 — 70 мВ, 2) наличием МДД, 3) близкой к пикообразному потенциалу формой ПД, 4) низкой амплитудой ПД — 30 — 50 мВ без явления риверсии (овершута).
Клетки миокарда обладают возбудимостью, но им не присущаавтоматия. В период диастолы мембранный потенциал покоя этих клеток стабилен, и его величина выше (80—90 мВ), чем в клетках водителей ритма. Потенциал действия в этих клетках возникает под влиянием возбуждения клеток водителей ритма, которое достигает кардиомиоцитов, вызывая деполяризацию их мембран
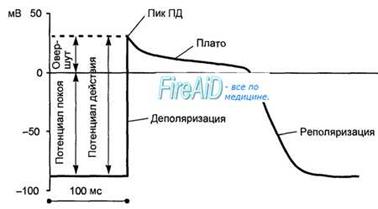
Потенциал действия клетки рабочего миокарда. Быстрое развитие деполяризации и продолжительнаяреполяризация. Замедленнаяреполяри-зация (плато) переходит в быструю реполяризацию. Потенциал действия клеток рабочего миокарда состоит из фазы быстрой деполяризации, начальной быстрой реполяризации, переходящей в фазу медленной реполяризации (фаза плато), и фазы быстрой конечной реполяризации (рис. 9.8). Фаза быстрой деполяризации создается резким повышением проницаемости мембраны для ионов натрия, что приводит к возникновению быстрого входящего натриевого тока. Последний, однако, при достижении мембранного потенциала 30—40 мВ инактивируется и в последующем, вплоть до инверсии потенциала (около +30 мВ) и в фазу «плато», ведущее значение имеют кальциевые ионные токи. Деполяризация мембраны вызывает активацию кальциевых каналов, в результате чего возникает дополнительный деполяризирующий входящий кальциевый ток 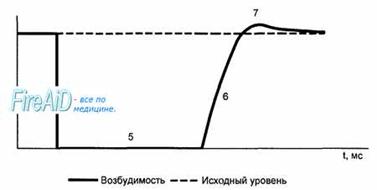
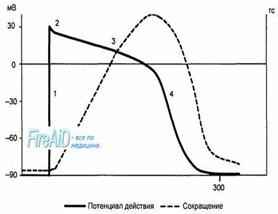
Рис. 9.9. Сопоставление потенциала действия и сокращения миокарда с фазами изменения возбудимости. 1 — фаза деполяризации; 2 — фаза начальной быстрой реполяризации; 3 — фаза медленной реполяризации (фаза плато); 4 — фаза конечной быстрой реполяризации; 5 — фаза абсолютной рефрактерности; 6 — фаза относительной рефрактерности; 7 — фаза супернормальной возбудимости. Рефрактерность миокарда практически совпадает не только с возбуждением, но и с периодом сокращения. Конечная реполяризация в клетках миокарда обусловлена постепенным уменьшением проницаемости мембраны для кальция и повышением проницаемости для калия. В результате входящий ток кальция уменьшается, а выходящий ток калия возрастает, что обеспечивает быстрое восстановление мембранного потенциала покоя. Длительность потенциала действия кардиомиоцитов составляет 300—400 мс, что соответствует длительности сокращения миокарда (рис. 9.9).

