 2013-12-31
2013-12-31 778
7781. Якія прыкметы раслін аддзела голанасенных?
2. Якія класы ўваходзяць у склад аддзела голанасенных?
3. Вызначце філагенетычнае становішча насенных папарацей.
4. Назавіце рысы прымітыўнай арганізацыі сагаўнікавых і гінкгавых.
5. Якія асаблівасці мае цыкл развіцця хвойных (на прыкладзе сасны звычайнай)?
6. Назавіце характэрныя прыкметы класа гнетавых.
АДДЗЕЛ МАГНОЛІЯФІТЫ, АБО ПАКРЫТАНАСЕННЫЯ
(MAGNOLIOPHYTA, ЦІ ANGIOSPERMAE)
Паходжанне магноліяфітаў
У батанікаў няма адзінай думкі пра паходжанне кветкавых раслін (магноляіфітаў).
А.Л. Тахтаджан лічыць, што кветкавыя расліны ўзніклі лакальна ў гарах Паўднёва-Усходняй Азіі (старажытны раён Катазія) у ніжнім – сярэднім меле (гл. табліцу 3).
Пад уздзеяннем механізмаў сінтэтычнай тэорыі эвалюцыі (СТЭ) – мутацый, зменлівасці, спадчыннасці, натуральнага адбору ў разнастайных умовах горнага рэльефу фарміраваліся розныя таксоны раслін пры чаргаванні перыядаў ізаляцыі і гібрыдызацыі, флюктуацыі колькасці раслін (папуляцыйныя хвалі). Пры змяненні колькасці і ізаляцыі зноў узніклая карысная прыкмета хутка асімілявалася геномам папуляцыі. Тахтаджан не выключае і “генетычны дрэйф”, або нязначныя змяненні формы, якія індыферэнтныя выконваемай функцыі. Лакальна ўзніклая разнастайнасць фарміравала фітацэнозы – мнагаярусныя супольніцтвы і “камплекты” – розныя варыянты расліннасці і флоры. Яны з адпаведных паясоў гор перамяшчаліся на шыротную занальнасць раўнін.
Такім чынам, па Тахтаджану, працяглы перыяд узнікнення разнастайнасці кветкавых раслін змяняўся даволі хуткім рассяленнем яе па тэрыторыі зямнога шара. Перамогу кветкавых раслін у барацьбе за існаванне Тахтаджан, услед за М.І. Галенкіным, тлумачыць іх лепшай адаптаванасцю да ўмоў змененага і больш сухога клімату, калі ў мелавым перыядзе мацерыкі прынялі сучасны выгляд і змяніліся суадносіны плошчаў сушы і мора.
Працэс узнікнення завязі кветкавых раслін шляхам загортвання спарафіла вакол семязачатка тлумачыцца Тахтаджанам як ахоўная рэакцыя на апыленне жукамі (кантарафілія). Пазней пад уплывам Н.В. Пярвухінай (1970 г.) ён стаў тлумачыць фарміраванне завязі неабходнасцю дадатковай аховы семязачаткаў ад высыхання.
Тахтаджан з’яўляецца прыхільнікам монафілетычнай гіпотэзы паходжання кветкавых раслін ад магноліепадобных старажытных продкаў (катазійная гіпотэза).
С.В. Меен на аснове аналізу палеабатанічных матэрыялаў паказаў, што кветкавыя расліны ўзніклі ў ніжнім меле ўздоўж усяго экватарыяльнага пояса, дзе спрыяльныя ўмовы дазвалялі выжываць і новым арганізмам з рэзкімі адхіленнямі формы (сальтацыі). У далейшым, у перыяды пацяплення трапічная “помпа” распаўсюджвала ўзніклыя формы ў больш высокія шыроты, дзе яны эвалюцыянавалі ўжо ў многім дзякуючы больш павольным адаптацыйным механізмам СТЭ. Пры гэтым узнікненне новых відаў звязваецца з любымі формамі рэльефу і не ўводзіцца ўяўленне аб фарміраванні фітацэнатычных “камплектаў” флоры, якія цалкам катапультаваліся з вертыкальных паясоў гор у адпаведныя зоны раўнін. Сучасныя фітацэналогія і экалогія разглядаюць эвалюцыю расліннасці як сеткападобны працэс і адмаўляюць магчымасць сур’ёзных каадаптацый арганізмаў аднаго трафічнага ўзроўню. У згуртаваннях яны сумесна ўтрымліваюцца механізмамі дыферэнцыяцыі ніш. Таму вялікую ролю могуць адыгрываць не толькі адаптацыі (прыстасаванні відаў да пэўных умоў нішы), але і абабтацыі, ці “эвалюцыі ад” – дывергенцыя патрэбнасцей відаў шляхам разыходжання іх па розных нішах. Віды ў гэтым выпадку становяцца непадобнымі адзін на аднаго па сваіх патрабаваннях, па патэнцыяльных і рэалізаваных (або адкарэктаваных канкурэнцыяй) нішах.
Меен у сваёй распрацоўцы гіпотэзы фітаспрэдзінгу (spread –распаўсюджваць) сыходзіў з палеабатанічных матэрыялаў. Да моманту фарміравання яго поглядаў (70-я гады) было ўстаноўлена, што на месцы Катазіі, дзе быццам адбываўся бурны прагрэс кветкавых у гарах (па меркаванні А.Л. Тахтаджана), наогул было мора. Сучасныя скапленні старажытных формаў кветкавых раслін у Паўднёва-Усходняй Азіі пацвярджаюць палеабатанічныя даныя аб тым, што старажытныя формы сустракаюцца практычна ўздоўж усяго экватара і іх таксанамічны склад толькі нагадвае сучасную флору, але не супадае з ёю.
Былі знойдзены і кветкі першых пакрытанасенных раслін, якія ніяк не адпавядаюць буйным кветкам сучасных магноліевых і з‘яўляюцца дастаткова несамавітымі і сабранымі ў суквецці. Гэта дазваляе лічыць больш правільным меркаванне Н.В. Пярвухінай (1970) аб тым, што першыя кветкавыя расліны не былі выключна насякомаапыляльнымі, а маглі апыляцца ветрам, насякомымі або самаапыляцца.
Слабымі месцамі абедзвюх гіпотэз з’яўляецца ўяўленне аб продках кветкавых раслін. Тахтаджан лічыць, што гэта былі насенныя папараці. Але нельга выключаць магчымасці ўзнікнення кветкавых ад іншых, яшчэ не вядомых прадстаўнікоў голанасенных. Няпаўната геалагічнага летапісу паступова знікае. Меен мяркуе, што продкамі кветкавых былі бенетыты, стробіл якіх быў двухполы. Але неадпаведнасць будовы стробіла бенетытаў з палачкападобнымі спарангіяфорамі і кветкі з лістападобнымі мегаспарафіламі, якія абгортваюцца вакол семязачатка, да канца не аргументавана даволі складанымі міграцыямі органаў унутры структуры “пракветкі”.
Антыноміі гіпотэз паходжання кветкавых раслін прыведзены ў табліцы 3 (Міркін, 1989).
В.М. Ціхаміраў (1987) лічыў, што ўзнікненне кветкі і пакрытанасеннасць адбываліся самым розным чынам і ўсю разнастайнасць у кветкавых нельга вывесці з якойсьці адной структуры.
Прыкметай, якая аб’ядноўвае ўсе варыянты будовы кветкі пакрытанасенных у адну групу, Ціхаміраў дапускаў наяўнасць працэсу двайнога апладнення. Гэтае меркаванне добра ўпісваецца ў агульную ідэю поліфілетычнасці, якую развівае С.В. Меен. Аднак і двайное апладненне так вар’іруе ў розных групах пакрытанасенных, што дапускае магчымасць узнікнення яго рознымі шляхамі.
Табліца 3 – Асноўныя антыноміі гіпотэз паходжання кветкавых раслін
| Пытанні | Катазійная гіпотэза Тахтаджана | Гіпотэза фітаспрэдзінгу Меена |
| Час паходжання | Ніжні і сярэдні мел | Ніжні – верхні мел |
| Час рассялення | Верхні мел | Ніжні – верхні мел |
| Месца | Паўднёва-Усходняя Азія | Трапічны пояс |
| Экалагічныя ўмовы | Горы | Розныя, але пераважна раўнінныя |
| Продкі | Насенныя папараці | Бенетытавыя |
| Старажытныя формы | Вечназялёныя дрэвы з адзіночнымі буйнымі кветкамі тыпу магноліевых | Розныя дрэвавыя расліны з несамавітымі кветкамі, сабранымі ў суквецці |
| Спосаб апылення | Энтамафілія, асабліва кантарафілія (апыленне жукамі) | Факультатыўная энтамафілія, анемафілія, самаапыленне |
| Асноўныя эвалюцыйныя механізмы ўзнікнення разнастайнасці | СТЭ. Адаптацыя да розных горных месцазнаходжанняў (па вышыні над узроўнем мора, экспазіцыі). Спалучэнне мутацый і накіраванага адбору. Папуляцыйныя хвалі. Генетычны дрэйф. Чаргаванне перыядаў ізаляцыі лакальных папуляцый (дэмаў) і гібрыдызацыі | Сальтацыі: выжыванне арганізмаў са значнымі змяненнямі формы неадаптыўнага характару, адпампоўванне іх “трапічнай помпай” у высокія шыроты і адаптацыйныя змяненні пад уплывам механізмаў СТЭ |
Такім чынам, пытанне пра паходжанне пакрытанасенных – іх мона-, ды-, поліфілетычнасці ўзнікнення – застаецца дыскусійным.
Агульная характарыстыка магноліяфітаў
Гэты аддзел вышэйшых раслін называюць таксама пакрытанасенныя (Angiospermae), кветкавыя (Anthophyta), песцічныя (Gynoeciatae). Ён уключае больш 450 сямей, каля 12,5 тыс. родаў і каля 250 тыс. відаў.
Па колькасці відаў пакрытанасенныя пераўзыходзяць усе астатнія групы вышэйшых раслін, узятыя разам. Кветкавыя расліны растуць ва ўсіх кліматычных зонах і ў самых разнастайных экалагічных умовах – ад трапічных лясоў да тундры, ад балот да пустынь і ад марскіх узбярэжжаў да высокіх гор. Яны складаюць асноўную масу расліннага рэчыва біясферы і з‘яўляюцца самай важнай для чалавека групай раслін. Усе важнейшыя культурныя расліны, у тым ліку хлебныя злакі і амаль усе агароднінныя расліны і пладовыя дрэвы, адносяцца да кветкавых раслін.
Ад усіх астатніх аддзелаў вышэйшых раслін, за выключэннем голанасенных, кветкавыя адрозніваюцца ўтварэннем насення. Прадстаўнікі абодвух гэтых аддзелаў з‘яўляюцца раслінамі насеннымі, у той час як усе астатнія аддзелы з‘яўляюцца безнасеннымі (рассяленне ў іх адбываецца пры дапамозе спор, а не насення). Ад голанасенных кветкавыя расліны адрозніваюцца перш за ўсё тым, што семязачаткі ў іх змяшчаюцца ў больш ці менш замкнутую поласць – завязь (утвораную адным ці некалькімі пладалісцікамі), з-за чаго іх часцей за ўсё называюць пакрытанасеннымі. Дзякуючы таму, што семязачаткі заключаны ў поласць завязі пылок трапляе не непасрэдна на мікропіле семязачатка, а на рыльца. Наяўнасць апошняга (якое ў прымітыўных форм цягнецца ўздоўж шва пладалісціка) з‘яўляецца характэрнейшай асаблівасцю кветкавых раслін і фактычна гэтым кветкі адрозніваюцца ад стробілаў голанасенных.
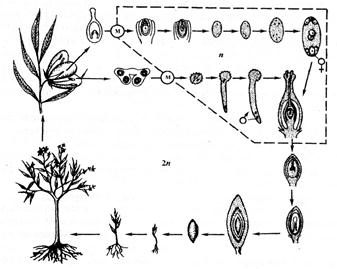
Гаметафіты (жаночы – зародкавы мяшок, мужчынскі – пылковае зерне) крайне спрошчаны і развіваюцца значна хутчэй, чым у голанасенных, у сувязі з чым яны страцілі гаметангіі – антэрыдыі і архегоніі.
Адной з характэрнейшых асаблівасцей кветкавых раслін з‘яўляецца так званае двайное апладненне, што рэзка адрознівае іх ад астатніх груп расліннага царства. Гэты працэс заключаецца ў тым, што адзін з двух утвораных у мужчынскім гаметафіце сперміяў (мужчынскіх гамет) зліваецца з яйцаклеткай (уласна апладненне), а другі – з двума свабоднымі ці ўжо зліўшыміся так званымі палярнымі ядрамі жаночага гаметафіта (трайное зліццё). У выніку зліцця аднаго са сперміяў з яйцаклеткай утвараецца зігота, а вынікам трайнога зліцця з‘яўляецца ўтварэнне характэрнай для кветкавых раслін пажыўнай тканкі для развіваючагася з зіготы зародка – трыплоіднага эндасперма. Насенне кветкавых раслін заключана ў плод (адсюль другая назва кветкавых раслін – пакрытанасенныя).
Цыкл развіцця пакрытанасенных раслин паказаны на малюнку 64.
У адрозненне ад большасці голанасенных для большасці кветкавых раслін характэрна наяўнасць у ксілеме побач з трахеідамі таксама сасудаў, якія адсутнічаюць толькі ў некаторых ці спецыяльных групах пакрытанасенных. Значныя адрозненні ёсць таксама ў будове флаэмы: сітападобныя трубкі кветкавых раслін забяспечаны так званымі клеткамі-спадарожніцамі, якія адсутнічаюць у голанасенных.
Усё гэта дазваляе сцвярджаць, што ў цэлым кветкавыя расліны дасягнулі больш высокага ўзроўню эвалюцыйнага развіцця, чым голанасенныя.
Параўнальнне будовы гаметафітаў, ходу палавога працэсу і развіцця семян у пакрытанасенных і голанасенных прыведзена ў табліцы 4.

Табліца 4 – Будова гаметафітаў, ход палавога працэсу, развіццё сямян ў пакрытанасенных і голанасенных раслін
| У пакрытанасенных | У голанасенных |
| 1. Жаночы гаметафіт, ці зарастак, у выглядзе 8-ядзернага зародкавага мяшка. | 1. Жаночы гаметафіт, ці зарастак, у выглядзе эндасперма (першаснага) з архегоніямі. |
| 4. Мужчынскі гаметафіт – прарослая пылінка з утварэннем спермагеннай клеткі і клеткі-трубкі, клетка трубка ўтварае пылковую трубку, спермагенная трубка- два спермія. | 4. Мужчынскі гаметафіт – прарослая мікраспора з утварэннем праталіяльных клетак і антэрыдыяльнай ініцыялі; антэрыдыяльная ініцыяль утварае антэрыдыяльную клетку і гаўстарыяльную клетку (клетку трубку); гаўстарыяльная клетка ўтварае гаўсторыю, клетка трбкі- пылковую трубку; антэрыдыяльная клетка ўтварае сперматагенную (спермагенную) клеткі і стэрыльную клетку; сперматагенная (спермагенная) клетка ўтварае або трох ядзер – пылковай трубкі і 2 сперматазоіды(сперміі). |
| 3. Двайное апладненне: адзін спермій апладняе яйцаклетку з утварэннем дыплоіднага зародка; другі – дыплоіднае ядро зародкавага мяшка, з утварэннем трыплоіднага другаснага эндасперма ў семені. | 3. Аднаразовае апладненне: адзін са сперміяў (сперматазоідаў) апладняе яйцаклетку аднаго з двух архегоніяў з утварэннем дыплоіднага зародка і гаплоідны першасны эндасперм у семені. |
| 4.Развіццё семязачатка і ўтварэнне насення адбываецца параўнальна хутка, асабліва ў травяністых форм, іншы раз за 3-4 тыдні (гранічна адзін вегетацыйны перыяд). | 4.Развіццё семязачаткаў і ўтварэнне насення адбываецца працягла (у сасны звычайнай каля 18 месяцаў), пры гэтым інтэрвал паміж апыленнем і апладненнем, напрыклад, у сасны каля – 13 месяцаў. |
| 5. Семязачатак знаходзіцца пад покрывам пладалісцікаў. | 5. Семязачатак размяшчаецца адкрыта,гола на насеннай лусцы. |
Агульнапрынятай гіпотэзы пра паходжанне і эвалюцыю кветкавых раслін (магноліяфітаў) няма (галоўным чынам з-за адсутнасці пераканаўчых выкапнёвых продкавых форм). Мноства агульных марфалагічных, анатамічных і эмбрыялагічных прыкмет (у тым ліку не звязаных паміж сабой у антагенезе і ў эвалюцыі) у самых розных сямей і парадкаў указвае на агульнасць паходжання ўсіх кветкавых раслін. Аднак ні адна з цяпер існуючых груп вышэйшых раслін не магла, як мяркуюць, даць пачатак кветкавым раслінам. Верагодна, іх продкі былі цесна звязаны з насеннымі папарацямі (птэрыдаспермамі) і вырасталі ў месцах, мала спрыяльных да адкладання і пахавання раслінных рэштак. Найбольш праўдападобна неатэнічнае паходжанне кветкавых раслін, асабліва паходжанне кветкі, а таксама жаночага і мужчынскага гаметафітаў. Хутчэй за ўсё, кветкавыя расліны ўзніклі ва ўмовах экалагічнага стрэсу, або ва ўмовах часовай засушлівасці мусоннага клімату, на адкрытых схілах, таму што неатэнія звычайна звязана з абмяжоўваючымі фактарамі асяроддзя. Верагодна, першаснымі кветкавымі раслінамі былі, драўняныя расліны з вечназялёнымі лістамі і двухполымі кветкамі ў прымітыўных цымозных (верхавінных) суквеццях.
Эвалюцыя кветкавых раслін ішла па шляху шырокай адаптацыі вельмі хуткімі тэмпамі. Самыя старажытныя выкапнёвыя рэшткі кветкавых раслін адносяцца да ранняга мелавога перыяду (каля 125 млн. гадоў назад). Дзякуючы высокай эвалюцыйнай пластычнасці кветкавыя расліны ў сярэдзіне мелавога перыяду (прыкладна 110 млн. гадоў назад) распаўсюдзіліся па ўсім зямным шары. Вялікую ролю ў эвалюцыі і рассяленні кветкавых раслін адыгралі насякомыя-апыляльнікі. Гэтыя расліны – адзіная група раслін, якая ўтварае складаныя шматярусныя супольнасці, што спрыяе больш інтэнсіўнаму выкарыстанню асяроддзя і больш паспяховаму заваяванню новых тэрыторый і асваенню новых месцавырастанняў.
Усю разнастайнасць форм росту кветкавых раслін можна звесці да двух асноўных тыпаў – драўнянаму і травяністаму. Драўняныя расліны характэрны для лясоў і куставых згуртаванняў, але адсутнічаюць ці выконваюць толькі другасную ролю ў стэпах, на лугах і іншых травяных згуртаваннях. Драўнінныя і асабліва травяністыя кветкавыя расліны характарызуюцца вялікай разнастайнасцю. Паміж драўніннымі і травяністымі раслінамі маюцца прамежкавыя формы. Для драўнінных раслін характэрна наяўнасць актыўна дзейнічаючага камбію. У травяністых раслін, як правіла, такі камбій адсутнічае ці маецца ў зачатачнай форме.
Лік відаў травяністых кветкавых раслін у цяперашні час значна перавышае колькасць відаў дрэў і кустоў. У працэсе эвалюцыі травы ўзніклі ад драўняных форм. Эвалюцыйнае ператварэнне драўняных форм ў травяністыя адбывалася ў выніку паступовага аслаблення і, нарэшце, поўнага спынення актыўнасці камбію. Падабенства анатамічнай будовы травяністых відаў тых родаў, дзе ёсць як травы, так і драўнінныя формы, у многім падобна на будову ювенільнай фазы роднаснага драўнянага віду. Гэта пацвярджае тое, што травы паходзяць ад драўняных форм у выніку неатэніі.
Нямногія травы ўзніклі ў выніку гідрафільнай эвалюцыі. Да воднага асяроддзя травы прыстасаваны лепш, чым кусты, а тым больш дрэвы.
Адным з важнейшых фактараў фарміравання травяністых форм было прыстасаванне да халоднага клімату з характэрным для яго кароткім вегетацыйным перыядам.
Асноўным органам размнажэння пакрытанасенных раслін з‘яўляецца кветка (ад лац. flos, грэч. anthos).
У двухполай кветцы адбываецца мікра- і мегаспарагенез, апыленне, апладненне, развіццё зарастка і ўтварэнне плода з насеннем. Кветкі надзвычай разнастайныя па будове, афарбоўцы і памерах ад некалькіх міліметраў да 1 м у дыяметры (у рафлезіі – Rafflesia). Кветка часта мае кветаножку (калі яе няма, кветка называецца сядзячай). Усе часткі кветкі сядзяць на восі – кветаложы. Вонкавыя часткі кветкі – чашалісцікі і пялёсткі, якія разам утвараюць калякветнік.
Чашалісцікі ў большасці выпадкаў служаць для аховы кветкі, а калі яны зялёныя, ці ўтрымліваюць хларафілавыя зярняты, дык таксама і дадаткова органамі фотасінтэзу. Чашалісцікі маюць відавочную ліставую прыроду, яны паходзяць ад верхавінных вегетатыўных лістоў.
Сукупнасць чашалісцікаў кветкі называеца чашачкай (ад лац. calyx). У працэсе эвалюцыі кветкі чашалісцікі часта зрастаюцца паміж сабою, утвараюць зрослалістую чашачку. У многіх выпадках у шчыльных густых суквецях чашачка рэдукуецца ў лётальны апарат.
Пялёсткі выконваюць іншую і прытым больш спецыялізаваную ролю, чым чашалісцікі. Іх асноўная функцыя – прывабліванне апыляльнікаў і садзеянне паспяховаму апыленню. Паходжанне пялёсткаў часцей за ўсё іншае, чым чашалісцікаў. Многія даследчыкі лічаць, што ў пераважнай большасці выпадкаў пялёсткі паходзяць ад тычынак. Прыводзіцца класічны прыклад ператварэння тычынак у пялёсткі прадстаўнікоў сям‘і гарлачыкавыя (Nyphaea alba і іншых), дзе назіраецца паступовы пераход ад тычынак да пялёсткаў. Тычыначная прырода пялёсткаў бачна ў прадстаўнікоў такіх сямей, як казяльцовыя, макавыя, гваздзіковыя, дыленіевыя, ружавыя, сусакавыя, шальніковыя і інш.
Пераўтварэнне тычынак у пялёсткі часта назіраецца ў выглядзе адхіленняў ад нармальнага развіцця, так званых выродлівасцей. У выніку ўзнікаюць махровыя формы кветак многіх дэкаратыўных раслін.
Аднак не ва ўсіх кветкавых раслін пялёсткі маюць бясспрэчна тычынкавае паходжанне. Напрыклад, у прадстаўнікоў сям‘і магноліевыя ці роду півоня пялёсткі звязаны ў сваім паходжанні з чашалісцікамі, а ў некаторых (вінтэравыя, бадзьянавыя, лімоннікавыя) ліставое паходжанне.
Сукупнасць пялёсткаў кветкі называюць венчыкам (ад лац. – corolla).Венчык адрозніваецца высокай эвалюцыйнай пластычнасцю і падвяргаецца вельмі вялікім марфалагічным зменам, звязаным з біялогіяй апылення. Таму памеры, будова і афарбоўка венчыка адрозніваецца вялікай разнастайнасцю. У той жа час у многіх кветкавых раслін венчыкі больш ці менш недаразвітыя або нават зусім адсутнічаюць, што звязана з прыстасаваннем да ветра-, самаапылення, або нават з пераходам іх функцыі да тычынак.
Як і чашалісцікі, пялёсткі часта зрастаюцца краямі, утвараюць зрослапялёсткавы венчык. Апошні ўзнік ад свабоднапялёсткавага незалежна ў розных лініях эвалюцыі кветкавых раслін.
Калі ў кветцы маюцца як чашачка, так і венчык, дык такі калякветнік называюць двайным. Калі ў кветцы пялёсткі адсутнічаюць ці калі няма добра выражаных адрозненняў паміж чашачкай і венчыкам, такі калякветнік называюць простым.
Тычынкі як па функцыі, так і па марфалогіі гамалагічныя мікраспарангіяфорам. У пераважнай большасці выпадкаў тычынка мае ніткападобную форму з адной жылкай. Аднак самыя прымітыўныя формы тычынак у такіх раслін, як дэгенерыя фіджыйская (Degeneria vitiensis) і шэрагу прадстаўнікоў магноліевых і вінтэравых, уяўляюць сабой адносна шырокую стужкападобную і звычайна трохнервовую пласцінку, мала падобную на ніць ці тычынку. Яны яшчэ не дыферэнцыраваны на тычыначную ніць, пыльнік, звязнік і, такім чынам, адрозненні паміж фертыльнай і стэрыльнай часткамі адсутнічаюць ці слаба выражаны. Такія архаічныя стужкападобныя тычынкі маюць чатыры папарна збліжаныя падоўжаныя лінейныя мікраспарангіі, больш ці менш пагружаныя ў тканку пласцінкі. Яны размешчаны на паверхні пласцінкі паміж сярэдняй жылкай і бакавымі жылкамі звычайна трохнервовых тычынак.
У працэсе эвалюцыі гэты больш старажытны тып тычынкі, які захаваўся толькі ў некаторых родаў, паступова дыферэнцыраваўся ў тычыначную нітку і пыльнік. Апошні складаецца з двух палавінак, злучаных паміж сабой звязнікам, які ўяўляе сабой працяг тычыначнай ніткі. Кожная палавінка пыльніка складаецца з двух мікраспарангіяў, звычайна называемых гнёздамі пыльніка, ці пылковымі мяшкамі. Па меры рэдукцыі і амаль знікнення звязніка, як у злакаў – дзве двухгняздовыя палавінкі пыльніка злучаюцца ў адзін чатырохгняздовы пыльнік. Калі поласці двух мікраспарангіяў пры паспяванні злучаюцца ў адну, пыльнік называецца аднагняздовым.
У некаторых спецыялізаваных кветках тычынкі губляюць асноўную і першапачатковую функцыі, становяцца стэрыльнымі і ператвараюцца ў так званыя стамінодыі, нектарнікі.
Сукупнасць тычынак у кветцы называецца андрацэем. У некаторых адносна больш прымітыўных сямей тычынкі размяшчаюцца па спіралі і лік іх не вызначаны, іншы раз даволі вялікі, але ў большасці кветкавых яны размешчаны цыклічна ці кальчакамі і лік іх пэўны, часцей за ўсё невялікі. Павелічэнне колькасці тычынак звязана з біялогіяй апылення і часцей за ўсё сустракаецца ў кветак без нектару. Аднак у эвалюцыі кветкавых раслін часцей адбываецца не павелічэнне, а памяншэнне ліку тычынак, і ў некаторых выпадках андрацэй складаецца з адной тычынкі.
У працэсе эвалюцыі андрацэя ніці тычынак часта зрастаюцца як паміж сабой, так і з іншымі членамі кветкі. Наогул як асобныя тычынкі, так і андрацэй у цэлым характарызуецца, як і іншыя часткі кветкі, высокай эвалюцыйнай пластычнасцю.
Унутраную частку кветкі, ці верхавінную вобласць кветаложа, займаюць пладалісцікі, ці карпелы.
Функцыянальна і марфалагічна пладалісцікі адпавядаюць мегаспарафілам. Адносна найбольш прымітыўны тып пладалісціка захаваўся ў роду дэгенерыя. У яго пладалісцік складаецца з кароткай ножкі, ці гінаподыя (gynopodium ад грэч. gyne – жанчына і pus (podus) – нага), і адносна тонкай пласцінкі, размешчанай уздоўж сярэдняй жылкі.
Калі разгарнуць пладалісцік дэгенерыі, то адкрыецца выемчатая на верхавіне лістападобная пласцінка, уздоўж якой праходзіць тры самастойныя жылкі (праводзячыя пучкі), сярэдняя (дарсальная) моцна разгаліноўваецца. Паміж сярэдняй жылкай і дзвюма бакавымі размешчаны шматлікія семязачаткі. Прымітыўнасць такіх пладалісцікаў падкрэсліваецца таксама тым, што некаторыя з іх не дасягнулі яшчэ поўнай замкнутасці. Пладалісцік дэгенерыі, краі якога, за выключэннем ніжняй яго часткі, не зрасліся, а толькі датыкаюцца. Пры гэтым уваход у пладалісцік закрываюць шматлікія кароткія залозістыя валаскі. Зрастанне гэтых збліжаных паверхней адбываецца толькі пасля цвіцення, у працэсе развіцця плода. Залозістыя валаскі ў дэгенерыі выконваюць і іншую ролю: яны не толькі перашкаджаюць уваходу ўнутр пладалісціка непажаданых наведвальнікаў, але сваімі выдзяленнямі спрыяюць успрыняццю пылку і яго прарастанню. Такім чынам, тут ужо маецца спецыяльная рыльцавая вобласць, якая ў дэгенерыі працягваецца шырокай палоскай па ўсёй даўжыні пладалісціка. Апошні ўтварае ўнутры поласці даволі шырокія палоскі, якія даходзяць амаль да семязачаткаў.
Па меры таго, як рыльцавая паверхня лакалізуецца ў верхняй частцы пладалісціка, з ніжняй часткі ўтвараецца фертыльная частка (завязь), над якой выцягваецца стэрыльнае слупковае ўтварэнне з рыльцам. Гэтыя спецыялізаваныя больш тонкія часткі пладалісціка называюцца слупком. Іншы раз слупок асобных пладалісцікаў называюць стылодыем.
Такім чынам, прымітыўны пладалісцік тыпу дэгенерыі паступова дыферэнцыруецца на завязь і слупок (стылодый). У працэсе далейшай эвалюцыі нізбягаючыя рыльцавыя грэбні паступова скарачаюцца і нарэшце рыльца лакалізуецца на верхавіне слупка.
Сукупнасць пладалісцікаў у кветцы называецца гінецэем (gynoecium ад грэч. gyne – жанчына і oikia – дом). Найбольш прымітыўны тып гінецэя характарызуецца свабоднымі пладалісцікамі, размешчанымі па спіралі. Гінецэй, утвораны свабоднымі пладалісцікамі, называецца апакарпным(ад грэч. apo – прыстаўка са значэннем адмаўлення, выдалення, страты і karpos – плод). Апакарпны гінецэй характэрны для адносна прымітыўных сямей.
У працэсе эвалюцыі пладалісцікі паступова зрасталіся паміж сабой і з апакарпнага гінецэя ўзнікае цэнакарпны гінецэй (ад грэч. kainos – агульны і karpos - плод. Цэнакарпны гінецэй бывае трох тыпаў: сінкарпны, паракарпны і лізікарпны.
Сінкарпным (ад грэч. syn – разам, і karpos - плод) называецца гінецэй утвораны некалькімі пладалісцікамі, зрослымі паміж сабой бакавымі сценкамі.
Паракарпны гінецэй (ад грэч. para – побач, каля і karpos - плод) –гініцэй утвораны некалькамі пладалісцікамі са зрослымі краямі і аднагняздовац завяззю.
Лізікарпны(ад грэч. lysis – развязванне, растварэнне і karpos - плод) гінецэй - гэта гінецэй утвораны некалькімі зрослымі пладалісцікамі. Іх бакавыя сценкі ў працэсе фарміравання гінецэя знікаюць, за выключэннем зрослых краёў, якія нясуць плацэнты, называемыя калонкай.
У кветкавых раслін адрозніваюць кветкі з верхняй завяззю, калі гінецэй размешчаецца вышэй за калякветнік; з ніжняй завяззю, калі гінецэй размяшчаецца ніжэй калякветніка, і паўніжняй, калі калякветнік адыходзіць ад сярэдзіны завязі.
Па сіметрыі кветкі падзяляюцца на правільныя, актынаморфныя, праз якія можна правесці некалькі восей сіметрыі, і на зігаморфныя, няправільныя кветкі з адной воссю сіметрыі.
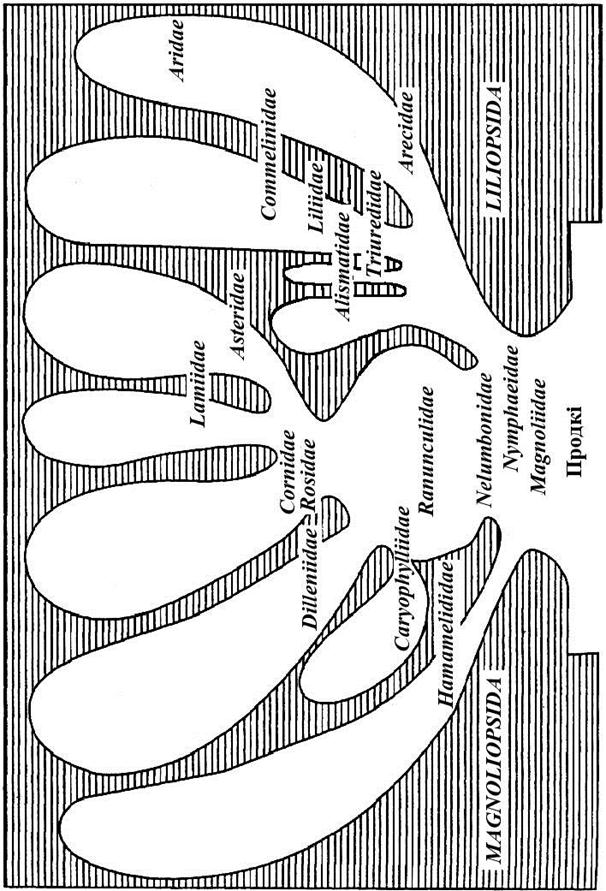
Агульнапрынятай філагенетычнай сістэмы пакрытанасенных няма. Рознагалоссі паміж аўтарамі сістэм пачынаюцца ўжо з зыходнага пункта – якія групы раслін неабходна лічыць больш прымітыўнымі і што прымаць для пакрытанасенных – мона-, бі- ці поліфілетычнае паходжанне.
Па схеме А. Энглера і Р. Ветштэйна (канец ХІХ – пачатак ХХ ст.ст.), найбольш прымітыўнымі сярод пакрытанасенных лічацца расліны з аднапокрыўнай ці бяспокрыўнай несамавітай кветкай (казуарыны, перцавыя, вярбовыя, бярозавыя, букавыя і г.д.).
Па сістэме Н. Буша, Б. Коза-Палянскага, А. Грасгейма, Г. Галіра, І. Бэсі, А. Тахтаджана і іншых батанікаў у яе пачатку знаходзяцца сем’і з добра развітымі мнагачленнымі, раздзельнапялёсткавымі, двухпокрыўнымі, энтамафільнымі кветкамі (магноліевыя, казяльцовыя, гарлачыкавыя і інш.).
Большасць сістэматыкаў дзеляць пакрытанасенныя на два класы: двухсемядольныя (двухдольныя – Magnoliopsida, ці Dicotyledones) і аднасемядольныя (аднадольныя – Liliopsida, ці Monocotyledones). Штучнасць такога дзялення відавочная. Англійскі батанік Э. Сарджэнт прыводзіць 40 агульных прыкмет для раслін абодвух класаў.
У вучэбным дапаможніку прынята дапоўненая філагенетычная сістэма кветкавых раслін (магноліяфітаў) па А.Л. Тахтаджану (малюнак 65).
Аддзел магноліяфіты падзяляецца на два класы – двухсемядольныя і аднасемядольныя.
Параўнальная характарыстыка адрозненняў класаў двух- і аднасемядольных прыводзіцца ў табліцы 5.

Табліца 5 – Асноўныя адрозненні паміж двухсемядольнымі і аднасемядольнымі
| Двухсемядольныя | Аднасемядольныя |
| Зародак семені звычайна з дзвюма семядолямі, якія, як правіла, прарастаюць надземна. Іншы раз зародак з адной семядоляй (напрыклад, у чыстацела – Ficaria, відаў чубаткі – Corydalis і некаторых парасонавых – Umbelliferae); рэдка зародак мае тры-чатыры семядолі (роды дэгенерыя – Degeneria і ідыяспермум – Idiospermum). Семядолі звычайна з трыма галоўнымі праводзячымі пучкамі. | Зародак семені з адной семядоляй, якая ў большасці выпадкаў прарастае падземна. Семядолі звычайна з двума галоўнымі праводзячымі пучкамі. |
| Лісты звычайна з перыстым ці, радзей, пальчатым, іншы раз дугавым ці паралельным жылкаваннем; звычайна маюцца свабодныя канцы жылак (жылкаванне незамкнутае). Чаранок звычайна добра выражаны, лісты рэдка маюць похвавую аснову. Ліставых слядоў, як правіла, налічваецца адзін-тры, іншы раз больш. | Лістызвычайна з так званым паралельным жылкаваннем ці з больш або менш паралельным размяшчэннем галоўных жылак, якія, аднак, злучаюцца паміж сабой кароткімі бакавымі жылкамі; радзей жылкаванне дугавое і вельмі рэдка пальчатае ці перыстае; свабодных канцоў жылак, як правіла, не бывае (жылкаванне звычайна замкнутае). Лісты звычайна не расчлененыя на чаранок і пласцінку, часта з похвавай асновай. Колькасць ліставых слядоў звычайна вялікая |
| Перадлісты(прафілы), ці самыя ніжнія, недаразвітыя лісты бакавых вегетатыўных парасткаў і адпаведныя ім прыкветнікі (брактэолі) звычайна парныя і размешчаны латэральна (за выключэннем некаторых прымітыў-ных груп, такіх, як анонавыя(Annonaceae), кірказонавыя, гарлачыкавыя (Nymphacaecae) і віды казяльца, у якіх яны няпарныя і размешчаны вентральна). | Перадлістыі брактэолі няпарныя (адзіночныя) і размешчаны на вентральным (брушным) баку парастка ці, радзей, парныя і размешчаны латэральна (жабнікавыя (Hydrocharitaceae), наядавыя, многія прадстаўнікі парадку лілейных і сямей асаковых і злакаў). |
Працяг таблiцы 5
| Праводзячая сістэма сцябла звычайна складаецца з аднаго кальца праводзячых пучкоў, як правіла, з камбіем (камбій адсутнічае ў некаторых сямей, у тым ліку ва ўсіх прадстаўнікоў парадку німфейных). У флаэме звычайна маецца парэнхіма (адсутнічае ў некаторых сямей, у тым ліку ў кірказонавых і казяльцовых). Кара і асяродак звычайна добра дыферэнцыраваны. | Праводзячая сістэма сцябла звычайна складаецца з многіх асобных пучкоў ці, іншы раз, з двух ці большага ліку кольцаў праводзячых пучкоў; праводзячыя пучкі звычайна пазбаўлены камбію (але рэшткавы пучковы камбій назіраецца ў прадстаўнікоў шэрагу сямей, у тым ліку ў роду красаднеў (Hemerocallis). У флаэме няма парэнхімы. Звычайна няма выразна дыферэнцыраваных кары і асяродка. |
| Першасны (зародкавы) карэньчык звычайна развіваецца ў галоўны корань, ад якога адыходзяць дробныя другасныя (бакавыя) карані (але ў многіх травяністых форм каранёвая сістэма валасніковістая). Чахольчык і эпідэрма маюць у антагенезе агульнае паходжанне (за выключэннем німфейных). | Першасны карэньчык адмірае, замяняецца сістэмай адвентыўных (прыдаткавых) каранёў, якія звычайна ўтвараюць валасніковістую каранёвую сістэму. Чахольчык і эпідэрма маюць у антагенезе рознае паходжанне. |
| Драўняныя ці травяністыя расліны (травяністыя расліны ўзнікаюць з драўняных). Іншы раз гэта другасныя дрэвападобныя формы, як, напрыклад, саксаул. | Звычайна травы, іншы раз другасныя дрэвападобныя формы (як, напрыклад, пальмы). Першасна драўняныя расліны адсутнічаюць. |
| Кветкі 5- ці, радзей, 4-членныя, і толькі ў некаторых, пераважна прымітыўных, груп бываюць 3-членныя (напрыклад, у анонавых і кірказонавых). | Кветкі звычайна 3-членныя, іншы раз 4- ці 2-членныя, але ніколі не бываюць 5-членнымі. |
| Нектарнікі розных тыпаў, часта ўяўляюць сабой відазмененыя тычынкі, рэдка бываюць септальнымі,ці размешчанных на перагародках завязі (ад лац. Septum - перагародка). | Нектарнікіпераважна септальныя, ці размешчаны на перагародках завязі |
| Пры дзяленні мацярынскай клеткі мікраспор клетачныя перагародкі закладваюцца пераважна па сімультанным тыпе. | Пры дзяленні мацярынскай клеткі мікраспор клетачныя перагародкі закладваюцца пераважна пасукцесіўным тыпе. |
Заканчэнне таблiцы 5
| Абалонка пылковых зярнят звычайна трохбарозная ці вытворных ад яе тыпаў (аднабарозная бывае толькі ў некаторых прымітыўных груп). | Абалонка пылковых зярнят звычайна аднабарозная ці вытворных ад яе тыпаў (часцей за ўсё аднапоравая), але ніколі не бывае 3-барознай. |
| Эндаспермцэлюлярны (ад лац. Cellula клетка), ці нуклеарны(ад лац.nucleus-ядро), рэдка гелабіяльны, ці базальны (прамежкавы). | Эндаспермгелабіяльны ці нуклеарны, вельмі рэдка цэлюлярны. |
Як відаць з табліцы, адрозненні паміж класамі кветкавых раслін у значнай ступені ўмоўныя. Але такое дзяленне пакрытанасенных раслін на класы дазваляе высветліць іх асаблівасці.








