 2014-02-02
2014-02-02 7602
7602Аллогенные сукцессии обусловлены влиянием внешнего фактора, который действует на фитоценоз. При этом чаще всего по мере ухудшения условий среды происходит смена видов в направлении их более высокой патиентности и приспособленности к фактору, вызвавшему сукцессию, то есть, такие сукцессии соответствуют модели толерантности. Реже аллогенные сукцессии протекают по модели благоприятствования, например, при удобрении лугов или эвтрофикации (повышении содержания в воде питательных элементов) водоемов. Но в этом случае уменьшается видовое богатство фитоценозов. В его отсутствие аллогенная сукцессия сменяется автогенной восстановительной.
При гологенезе, как уже говорилось, вследствие обусловленных различными причинами процессов развития ландшафтов сопряженно изменяется растительность целого ландшафта или его существенной части. Такие сукцессии могут быть природными и антропогенными.
| Рис. 32. Зоны речной поймы средней полосы европейской части России: П – пионерных сообществ, И – ивы, Т – тополя, В – вяза, Л – липы, О – черной ольхи; А-Б - поперечный профиль (Миркин и др., 2002). |
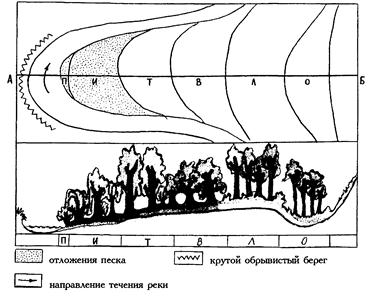 Как пример природной гологенетической сукцессии можно рассматривать процессы изменения растительности, происходящие при развитии речной долины (рис. 32). С одной стороны, текущая в реке вода постоянно размывает и углубляет ее русло, с другой стороны, поверхность поймы повышается за счет отложения взвешенных частиц, переносимых водой во время паводка (так называемого наилка). Кроме того, при меандрировании с одной стороны русла реки постоянно разрушается часть высокого берега, а с противоположной – наращивается песчаный пляж. В результате вышеописанных процессов на вновь образующихся намываемых субстратах происходит смена растительности по ряду: пионерные группировки – ивняк – осинник – вязовый лес – липово-дубовый лес. Кроме того, к террасе будет примыкать притеррасная пойма – область выклинивания грунтовых вод, где формируются низинные болота из Alnus glutinosa.
Как пример природной гологенетической сукцессии можно рассматривать процессы изменения растительности, происходящие при развитии речной долины (рис. 32). С одной стороны, текущая в реке вода постоянно размывает и углубляет ее русло, с другой стороны, поверхность поймы повышается за счет отложения взвешенных частиц, переносимых водой во время паводка (так называемого наилка). Кроме того, при меандрировании с одной стороны русла реки постоянно разрушается часть высокого берега, а с противоположной – наращивается песчаный пляж. В результате вышеописанных процессов на вновь образующихся намываемых субстратах происходит смена растительности по ряду: пионерные группировки – ивняк – осинник – вязовый лес – липово-дубовый лес. Кроме того, к террасе будет примыкать притеррасная пойма – область выклинивания грунтовых вод, где формируются низинные болота из Alnus glutinosa.
Вырубая леса в границах водосбора, человек может ускорять эту сукцессию. Вырубки, как правило, приводят к усиленной эрозии почвы, вследствие чего в пойму с весенними водами будет поступать большее количество смытой почвы. Это приведет к тому, что паводки станут более высокими, а процесс разрушения и созидания поймы активизируется. Облесением же водосбора человек может, напротив, снизить скорость процесса развития речной долины.
Как пример антропогенного гологенеза могут рассматриваться изменения растительности, происходящие в результате постройки крупных гидротехнических сооружений, например водохранилищ. Во первых, в результате затопления водами водохранилища наземных фитоценозов на их месте формируются водные сообщества. Во-вторых, выше плотины повышается уровень грунтовых вод и происходит подтопление почвы. Это приводит к тому, что существовавшие ранее на данной территории леса и травяная растительность сменяются фитоценозами, устойчивыми к высокому уровню грунтовых вод. Так, к примеру, вместо липово-дубовых, еловых или сосновых лесов формируются ольшаники, а вместо сухих лугов – осоковые болота. Ниже же плотины, наоборот, происходит понижение уровня грунтовых вод и, как следствие этого, иссушение поймы, а в южных районах – еще и засоление почв. В третьих, водохранилище изменяет климат, который становится более мягким и более ровным.
Оба рассмотренных примера показывают основную особенность гологенеза: под влиянием внешних факторов согласованно и закономерно изменяется совокупность фитоценозов на обширных территориях.
Гейтогенетические сукцессии – это изменения фитоценозов под влиянием действия внешних факторов, которые не связаны с общими тенденциями развития ландшафта. Как правило, основным фактором, вызывающим гейтогенетические сукцессии, является человек. В результате его деятельности происходят, главным образом, регрессивные сукцессии, называемые дигрессиями. Считается, что если в фитоценозе сохранилась хотя бы половина видов, то после прекращения действия фактора, вызвавшего дигрессивные изменения, возможно самовосстановление фитоценоза в результате процесса обратной трансформации, то есть вторичной автогенной сукцессии, которая называется восстановительной, или демутацией.
Рассмотрим некоторые примеры подобных сукцессий.
Сукцессии под влиянием выпаса ( пастбищные дигрессии). Это самый распространенный вариант сукцессии. Перевыпас и, как следствие, происходящее при этом обеднение фитоценозов связаны с избыточным поголовьем скота. На сегодняшний день подобные сукцессии являются одним из самых тяжелых последствий хозяйственной деятельности человека.
Выпас действует на травяные фитоценозы двояко. С одной стороны, разные виды растений избирательно поедаются скотом, что в итоге приводит к изменению видового состава пастбищ. С другой стороны, чрезмерный выпас скота приводит к уплотнению почвы и изменению ее водного, воздушного и солевого режимов. В результате высокотравные и малоустойчивые к выпасу растения сменяются пастбищным низкотравьем: подорожником средним, клевером ползучим, одуванчиками, лапчаткой гусиной и др. При этом уменьшается видовое богатство фитоценоза, упрощается структура, снижается общее проективное покрытие.
Рассмотрим стадии пастбищной дигрессии.
1. Стадия недостаточного выпаса. Для этой стадии, когда пастбищная нагрузка на фитоценоз очень слаба или вообще отсутствует, характерно скопление большого количества отмерших надземных органов растений, что препятствует нормальному росту типичных луговых трав. Семенное размножение дерновинных злаков подавлено, места, освобождающиеся после отмирания старых дерновин, занимают главным образом корневищные злаки. Поскольку испарение с поверхности почвы из-за накопления опада уменьшается, происходит мезофитизация травостоя.
2. Стадия умеренного выпаса. Все дерновинные злаки хорошо развиты, много их молодых дерновин, что говорит об их успешном семенном размножении. Одно- и двулетние виды подавлены. В этой стадии луг дает максимальную продукцию.
3. Стадия чрезмерного выпаса. Начинают исчезать или очень сильно подавляться ценные в кормовом отношении виды растений. Вместо их в зависимости от условий окружающей среды начинают преобладать практически не поедаемые скотом виды: осоки и щучка на сырых лугах, крупные щавели на богатых почвах, либо полыни на сухих почвах.
4. Стадия полного сбоя, или выгона. Из-за сильного механического разрушения дерновины копытами скота начинают преобладать однолетники со стелющимися побегами – спорыши, мятлик однолетний, дивалы и др.
В результате пастбищной дигрессии продуктивность травостоев может снижаться в десятки раз. Воспрепятствовать столь нежелательным последствиям можно лишь внедрением пастбищеоборота, при котором нагрузка скота на пастбище нормируется и разным участкам периодически предоставляется отдых.
Рекреационные сукцессии – это сукцессии, протекающие в растительности под влиянием специфической деятельности человека в процессе отдыха: вытаптывание, выжигание, засорение фитоценозов и т.д. Основным фактором, влияющим на фитоценоз в данном случае, является вытаптывание. Поэтому рекреационная сукцессия будет отличаться от пастбищной, главным образом, тем, что в данном случае отбор будет идти на устойчивые к вытаптыванию виды без учета их кормовых качеств.
Я. П. Дидух и П. Г. Плюта (1994) предлагают выделять следующие уровни рекреационной нагрузки:
1. рекреация отсутствует;
2. хотя и имеется определенная рекреационная нагрузка, но тем не менее сообщество еще сохраняет целостность своей структуры;
3. происходит смена доминирующих видов;
4. интенсивная рекреационная нагрузка ведет к локальному исчезновению растительного покрова, что дает возможность появиться в значительном количестве однолетникам, чаще всего эфемерам;
5. сбои, отсутствие травяного покрова из-за сильного вытаптывания.
В качестве примера можно рассмотреть антропогенную дигрессию в лесных сообществах. В лесах на ранних стадиях рекреационной сукцессии видовое богатство фитоценозов вначале повышается, так как лесные травы замещаются более устойчивыми к вытаптыванию луговыми растениями. Однако при усилении вытаптывания видовое разнообразие падает, начинают преобладать растения розеточных форм, устойчивые к вытаптыванию. В дальнейшем сукцессия захватывает подлесок и даже древесный ярус, так как уплотнение почвы препятствует возобновлению деревьев, ухудшает их состояние и даже может привести к усыханию древостоя.
Сукцессии под влиянием радиации. Эти сукцессии изучали Г. Вудвелл и Р. Уиттекер на экспериментальном полигоне на острове Лонг-Айленд (США). Результаты их исследований показали, что с повышением радиационной нагрузки, задаваемой гамма-излучением, изменения растительности происходят в обратной последовательности к сукцессии по модели благоприятствования при зарастании скал. Вначале исчезают деревья, потом кустарники, затем травы, мхи и лишайники.
После катастрофы на Чернобыльской АЭС в окрестностях взорвавшегося реактора также произошло усыхание деревьев (эту зону назвали "бурым лесом"), но кустарники и травы сохранились.
Внешние воздействия, вызывающие аллогенные сукцессии, могут воздействовать на фитоценозы как прямо, так и косвенно. В последнем случае в роли посредников между абиотическими факторами и растениями могут выступать как автотрофные, так и гетеротрофные компоненты экосистемы. В качестве примера подобного опосредованного влияния можно рассмотреть влияние так называемых "кислых" дождей, когда с дождем в почву вносится довольно значительное количество связанного азота, на вересковые пустоши Шотландии. В результате проведенных исследований оказалось, что исчезновение из растительных сообществ вереска при воздействии «кислых» дождей вызывается, по меньшей мере, двумя факторами. Во-первых, оно связано с увеличением конкурентной способности нитрофилов (как правило, рудеральных заносных видов), во-вторых, с тем, что вереск, активно поглощая азот, начинает сильно повреждаться фитофагами.
Говоря о сукцессиях, нельзя не упомянуть о таком понятии, как "климакс" в растительности. Концепция климакса (устойчивого состояния, к которому "стремится" сообщество в ходе автогенной сукцессии) разрабатывалась американскими экологами еще в конце 19-го – начале 20-го столетия. Наиболее полно она была развита Ф. Клементсом, представления которого получили название моноклимакса в растительности.
Суть взглядов Ф. Клементса сводилась к следующему. В пределах одной климатической зоны все фитоценозы в ходе сукцессии должны конвергировать к одному климаксовому сообществу. Например, в умеренном климате при достижении определенной мощности торфяной залежи болота должны зарасти лесом, на скалах после образования почвы также должен вырасти лес и т.д. При этом в каждой климатической зоне в результате сукцессии все фитоценозы должны сойтись в один тип. Для тайги это будет еловый лес, для степной зоны – ковыльная степь, для тундр – сообщества мелких ив, осок и лишайников. Причем конечный результат данного процесса не будет зависеть от типа субстрата, на котором будут формироваться фитоценозы: на скалах, песках, глинах, торфянистых почвах в конечном итоге сформируются растительные сообщества одного состава, с одинаковой структурой и функциональными связями.
Ф. Клементс считал, что климаксовый фитоценоз формируется очень долго – тысячи и десятки тысяч лет, причем допускалась возможность различных отклонений от основного климакса, который назывался климатическим. Так, сообщества, возникающие в результате постоянного действия человека, препятствующего формированию климакса (например, выпас), назывались дисклимаксовыми. Если же формированию климакса мешали местные экологические факторы (например, паводки в пойме реки), то образованные в этом случае фитоценозы назывались субклимаксовыми и т.д. К концу жизни (Клементс публиковал свои работы в течение почти пятидесяти лет) он выделил около тридцати различных вариантов климакса. Климаксовые устойчивые фитоценозы Ф. Клементс противопоставлял серийным, т. е. сукцессионным. Смена серийных фитоценозов приводит к климаксу.
Представления Ф. Клементса о моноклимаксе стали предметом вполне справедливой критики. В результате была сформулирована концепция поликлимакса: в одной климатической зоне фитоценозы различных местообитаний изменяются в ходе сукцессии, но в один тип не конвергируют, а на песках, скалах, болотистых местообитаниях и т.д. формируются свои климаксовые сообщества.
В 50-х годах, когда растительный покров начал рассматриваться как непрерывное явление, Р. Уиттекером был предложен третий вариант концепции климакса – концепция климакс - континуума. Главной чертой, принципиально отличающей ее от других концепций, явилась констатация того, что между климаксовыми фитоценозами существуют переходы, и поэтому количество климаксов в поликлимаксе стремится к бесконечности. При этом в каждой точке под влиянием уникального сочетания факторов формируется свой индивидуальный климакс.
Р. Уиттекеру также принадлежит оригинальная классификация климаксов, в качестве основного критерия которой рассматривается длительность жизни доминантов серийных и климаксовых сообществ.
· Аклимакс. В данном случае время генерации доминантов меньше цикла изменений условий среды и флуктуации сообществ постоянны. Климаксовые и серийные сообщества в таком случае неразличимы. Примером аклимакса могут служить сообщества планктона.
· Циклоклимакс. Такой тип устойчивого состояния сообщества, в котором время генерации доминантов совпадает с годичными колебаниями условий среды. Как климаксовые сообщества, так и серийные в данном случае могут быть охарактеризованы главными доминантами-однолетниками. В качестве примера циклоклимаксовых сообществ могут рассматриваться некоторые сообщества пустынь, формируемые однолетними растениями.
· Катаклимакс. В данном случае генерация доминантов происходит в период между повторяющимися нарушениями среды, уничтожающими растительность. Это могут быть, к примеру, пожары, сели. Климаксовые и серийные сообщества в данном случае будут в большей или меньшей степени различаться.
· Суперклимакс, наблюдающийся, например, в тундре. В данных условиях формируется крайне низкая биомасса сообщества и более или менее стабильные популяции растений. При этом генерация доминантов длительна, а изменения среды незначительны. Все это приводит к тому, что серийные и климаксовые сообщества формируются из одних и тех же видов и различаются лишь своей структурой.
· Эуклимакс. По-видимому, наиболее широко распространенное устойчивое состояние фитоценоза, возникающее в условиях длительной генерации доминантов и более или менее непрерывно изменяющегося состава доминантов в ходе сукцессии. Данные фитоценозы характеризуются, как правило, высокой биомассой. Серийные сообщества при этом хорошо отличаются от климаксовых. Примером подобного процесса может служить классическая "клементсовская" сукцессия, которая приводит к развитию лесной растительности.
В современной фитоценологии концепция климакса играет важную роль, но климакс не абсолютизируется, а понимается как тенденция формирования фитоценозов зонального типа.
В заключение следует еще раз обратить внимание на то, что флуктуации, автогенные и аллогенные сукцессии не происходят независимо друг от друга, а представляют единый процесс изменения растительных сообществ. Это связано, в первую очередь, с тем, что лежащие в их основе процессы неотделимы друг от друга. Флуктуации, автогенные и аллогенные сукцессии есть составляющие непрерывного процесса изменения растительности. Различаются же они, главным образом, по степени обратимости изменений (флуктуации) и по преобладающим причинам, заключенным либо в самих сообществах (автогенные сукцессии), либо вне их (аллогенные сукцессии). Но всегда следует помнить, что, во-первых, при флуктуации не может быть полной обратимости, так как не прекращается автогенная сукцессия, в то же время в ходе аллогенных сукцессий всегда наблюдаются эндогенетические процессы.
Классификация растительности (синтаксономия)
Любая отрасль знаний не может обойтись без упорядочения изучаемых объектов, их сортировки по тем или иным признакам, т. е. их классификации. Классификация является необходимым этапом любого исследования, так как без классификации, хотя бы предварительной, с одной стороны – трудно ориентироваться в материале и правильно планировать работу, а с другой стороны – классификация очень важна при завершении исследования, когда подводятся итоги сделанного.
Актуальность и необходимость классификации растительности связана с тем, что без данной процедуры невозможно обойтись во многих случаях: при описании растительного покрова, производимого с теми или иными целями и для любого региона; при выработке типологии хозяйственных угодий (лесных, сенокосных, пастбищных, рекреационных и др.); при сельскохозяйственной оценке земель и т.д. Большое значение имеет также правильное решение вопросов классификации при разработке приемов рациональной эксплуатации, улучшения и преобразования растительных сообществ. Полученные экспериментальным путем, эти приемы затем рекомендуются для определенных типов сообществ, и необходимо обеспечить правильность и единообразие типологии, чтобы апробированные рекомендации применялись для тех типов, для которых они вырабатывались и предназначались. Да и любые данные, полученные при изучении растительности, должны быть всегда отнесены к тому или иному типу растительных сообществ, изучается ли продуктивность растительных сообществ, их структура или взаимоотношения между его компонентами, связи со средой, сезонная или погодичная изменчивость и т.д., так как лишь при таком условии эти данные могут быть правильно интерпретированы и использованы в дальнейшей работе. Отсюда ясно, насколько важно, чтобы принципы и методы классификации были достаточно четко разработаны.
В фитоценологии, изучающей растительный покров с точки зрения слагающих его растительных сообществ, классификация (синтаксономия) в очень сильной степени отличается от классификации отдельных организмов (таксономии). Если организмы по природе своей дискретны, то есть отграничены в пространстве и во времени (смертны), то к фитоценозам вообще неприменимы такие понятия, как "особь", "индивид" и др. Дело в том, что растительное сообщество отличается низкой степенью целостности и регуляция в нем является статистическим результатом случайного поведения отдельных компонентов сообщества. Кроме того, как мы уже не один раз говорили, растительность сочетает в себе два противоположных свойства – дискретность и непрерывность. И именно непрерывность растительности создает основные проблемы при классификации: при любой классификации непрерывность приходится сводить до дискретности, то есть разбивать растительный покров на условно однородные участки, которые зачастую (но далеко не всегда!) переходят один в другой. Но это не значит, что если растительность непрерывна и растительные сообщества более или менее постепенно переходят одно в другое, то их нельзя классифицировать. Если проводить аналогии с какими-либо бытовыми вещами, то растительный покров можно сравнить со звуковым рядом или со световым спектром. К примеру, еще никто не утверждал, что нельзя пользоваться названиями оттенков цвета только потому, что они являются субъективно различаемыми частями непрерывного светового спектра.
Типы сообществ, которые выделяются в процессе классификации (синтаксоны), – это абстрактные классы, в каждом из которых сгруппировано определенное количество сообществ на основе присущих им свойств. Сообщества можно классифицировать по целому ряду признаков: жизненной форме доминанта, видам-доминантам, ярусной структуре, видовому составу и т.д. Различные характеристики сообщества, которые используются для классификации, не изменяются параллельно друг другу. Использование разных признаков одних и тех же сообществ приводит к разным классификациям, к выделению различных типов сообществ. В любой классификации границы между типами сообществ более или менее произвольны, поскольку они определяются признаками, взятыми исследователем в качестве критерия систематизации, и выбором способа проведения границ между типами. Не существует какого-либо единственно правильного способа разграничения сообществ. Поэтому были разработаны несколько отличных друг от друга систем классификации.
Если остановиться на методических подходах к классификации растительности, то, в первую очередь, следует различать два основных способа классификации: индуктивный и дедуктивный.
При дедуктивном способе классификация осуществляется "сверху", то есть путем деления имеющегося множества объектов на все более мелкие классы. При этом исследователю с самого начала должно быть известно все разнообразие объектов и их признаки, из которых он может выбрать те или иные для осуществления классификации. Примерами классификаций, произведенных дедуктивным способом, являются многочисленные физиогномические классификации растительности. В очень большой степени дедуктивным является геоботаническое районирование планеты и отдельных ее регионов.
При индуктивном подходе классификационная схема начинает строиться с низших таксономических единиц. В этом случае исследователь заранее не знает тех признаков, которые он положит в основу выделения тех или иных таксонов. Эти сведения он постепенно накапливает при последовательном изучении единичных объектов и, в результате, сообщества объединяются во все более крупные классы по степени сходства. В качестве примеров индуктивных способов классификации в геоботанике можно привести классификации Браун-Бланке, количественные методы классификации и др.
Перед тем, как перейти к системам классификаций растительности, в том числе основанным на индуктивном подходе, следует остановиться на основных единицах синтаксономии. Дело в том, что если при дедуктивном подходе к классификации мы работаем сразу со всем массивом имеющейся информации, постепенно разделяя его на части, то при индуктивном подходе мы, наоборот, должны иметь наименьшие единицы, чтобы их компоновать по степени сходства во все более крупные классы. Именно поэтому при подобных вариантах классификации должны быть основные унифицированные единицы классификации, с которыми мы будем работать.
Такая основная единица в фитоценологии – растительная ассоциация, обычно называемая просто ассоциацией. Ассоциация – это типологическая единица, тип фитоценоза. Если каждый фитоценоз конкретен, то ассоциация в одно и то же время, как указывает П. Д, Ярошенко (1962), "в некоторой мере абстрактна как тип фитоценозов и в то же время конкретна как их совокупность". Каждый фитоценоз, относящийся к той или иной ассоциации, наряду с общими, свойственными всем сходным фитоценозам признакам, может обладать и некоторыми случайными, индивидуальными, которые при характеристике ассоциации отбрасываются.
На Брюссельском ботаническом конгрессе в 1910 г. было предложено считать ассоциацию основной единицей растительного покрова. Принятое конгрессом определение этого термина таково: "Ассоциация есть растительное сообщество определенного флористического состава с особыми условиями местообитания, особой физиономией". Но данное определение в достаточной степени нечетко, поскольку ничего не говорится о том, какова должна быть степень однородности флористического состава, что такое особые условия местообитания и особая физиономия (облик сообщества). В конечном итоге эта нечеткость определения привела к тому, что ученые различных геоботанических школ при использовании различных систем классификации растительности по-разному понимали объем ассоциаций и выделяли их на основании разных критериев.
Так, среди советских геоботаников, придерживающихся доминантного подхода к классификации растительности, наиболее четко формулирует основные особенности ассоциации В. Н. Сукачев, который в 1938 г. писал: "…растительная ассоциация, или тип фитоценоза, объединяет фитоценозы, характеризующиеся однородным составом, строением и в основном одинаковым сложением составляющих их синузий и имеющие одинаковый характер взаимоотношений как между растениями, так и между ними и средой". То есть, ассоциация при такой трактовке должна включать фитоценозы, сходные по своей структуре, флористическому составу, доминантам в каждом ярусе, сезонной и разногодичной динамике и по условиям существования.








