 2015-01-30
2015-01-30 2289
2289Пектинметилэстеразы, ПЭ (К.Ф.3.1.11) катализируютотщепление метильных групп от полиметилгалактуроновой кислоты с образованиемметанола и частично деметоксилированной полигалактуроновой кислоты.
Процесс протекает согласно следующей схеме (стрелками показано действие фермента):
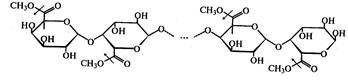
ПЭ деэтерифицирует пектины на 60-70%. По мере снижения степени этерификации субстратов уменьшается сродство фермента к ним, и процесс гидролиза не проходит до конца. ПЭ предпочтительно действует на крупные молекулы, метоксилированные олигоурониды расщепляются медленнее.
Пектинэстеразы синтезируются высшими растениями, микроскопическими грибами, дрожжами и бактериями.
Пектинэстеразы проявляют максимальную активность в интервале рН 4,4-8,0, у некоторых микроскопических грибов при рН 2,5. Оптимальная температура действия 30-40° С. Пектиностераза входит в комплексы пектолитических ферментных препаратов микробного происхождения.
Желирующая способность пектина зависит от степени метоксилиро-вания или степени этерификации, поэтому действие пектинэстеразы по отщеплению метоксильных групп приводит к снижению желирующей способности и сопровождается падением вязкости. На этом, очевидно, и основывается применение этого фермента для осветления плодовых соков и вина.
Обычно комплексные препараты пектолитических ферментов применяемые для этих целей, получают из различных плесневых грибов, и прежде всего из A. Niger
2.2. Гликозидазы (К.Ф.3.2)
Этот подкласс включает около ста ферментов с разной специфичностью действия, осуществляющих гидролиз олиго- и полисахаридов; некоторые ферменты этого типа способны осуществлять трансферазные реакции – переносит гликозидные остатки на олиго- и полисахариды, наращивать полисахаридные цепочки. Представители гликозидаз были одними из первых ферментов, обратимость действия которых in vitro была экспериментально доказана.
Гликозидазы стереоспецифичны. Они гидролизуют гликозидные связи определенной пространственной конфигурации (α или β), но не обеих одновременно. Примерами могут служить α-амилаза, α-глюкозидаза, глюкоамилаза, гидролизующие исключительно α-гликозидные связи, и β-глюканаза, целлобиза, лизоцим – гидролизующие β-связи.
Менее строгая избирательность проявляется к различным видам α- или β-гликозидных связей. Так, глюкоамилаза расщепляет α-1,4 и α-1,6-связи; глюканаза бацилл — β-1,3 и β-1,4-связи;
Гликозидазы специфичны в отношении длины цепи гидролизуемых полимерных субстратов. Так, глюкоамилаза плесневых грибов предпочтительно гидролизует высокомолекулярные полимеры α-1,4-связанной глюкозы, а аналогичный фермент – α-глюкозидаза дрожжей катализирует гидролиз той же связи в олигосахаридах, но не в крахмале.
Эндополигалактуроназа (К.Ф.3.2.1.15) и Экзополигалактуроназа (К.Ф.3.2.1.67)
Эти ферменты, наряду с пектинэстеразой осуществляют гидролиз пектиновых веществ.
Эндополигалактуроназа, эндо-ПГ (К.Ф.3.2.1.15) катализирует гидролиз α-1,4-гликозидных связей между неэстерифицированными остатками галактуроновой кислоты в различных пектиновых полисахаридах. Гидролиз происходит неупорядоченным способом, предпочтительно расщепляются внутренние связи полимеров. При гидролизе 1-10% связей вязкость растворов галактуронанов снижается не менее чем на 50%
|
 Полигалактуроновая кислота + Н20
Полигалактуроновая кислота + Н20
 Олигогалактуроновые кислоты + моногалактуроновая кислота
Олигогалактуроновые кислоты + моногалактуроновая кислота
Фермент расщепляет в пектинах фрагменты полигалактуроновой кислоты различной молекулярной массы и степени этерификации. С наибольшей скоростью гидролизуются низкоэтерифицированные субстраты, такие как пектовая кислота, полипектат натрия, свекловичный пектин. Более медленно - лимонный и яблочный пектин, а пектин Линка (степень этерификации 99%) совсем не гидролизуется. При действии на олигоурониды предпочтительно расщепляются субстраты большей степени полимеризации. Основным конечным продуктом гидролиза пектинов, в зависимости от источника фермента, может быть моно-, ди-, три- или тетрагалактуроновая кислота.
Действие эндо-ПГ на растительные ткани вызывает их мацерацию в результате расщепления пектиновых веществ срединных пластинок. В комплексе полигалактуроназ гриба Asp. alliaceus только эндо-ПГ практически полностью мацерировала до состояния изолированных клеток ткани картофеля, кабачка, тыквы, яблока, сливы, груши, персика.
Большинство исследованных эндополигалактуроназ – ферменты молекулярной массы 30-80 кДа с оптимумом действия при рН от 3,5 до 6,5 и температуре около 50° С. Эндо-ПГ некоторых микроскопических и высших грибов ингибируют ионы щелочноземельных и тяжелых металлов.
Экзополигалактуроназа экзо-ПГ (К.Ф.3.2.1.67) катализирует расщепление конечных α-1,4-глюкозидных связей между остатками неэстерифицированной галактуроновой кислоты в различных пектиновых полисахаридах (полигалактуроновой кислоте, пектатах, пектинах) с образованием моногалактуроновой кислоты. Гидролиз сопровождается незначительным снижением вязкости растворов субстратов. Фермент расщепляет пектаты неполностью, примерно на 50%.
Частично деградированные субстраты, как правило, гидролизуются экзо-ПГ быстрее высокополимерных. Предпочтительна низкая степень метоксилирования галактуронанов. Так, относительная скорость гидролиза субстратов разной степени этерификации двумя формами экзо-ПГ Asp. alliaceus составила: пектовой кислоты – 98-100, свекловичного пектина – 92-100, лимонного – 20-24, яблочного 1,3-7,5, сливового - 0,3-2,0.
Экзополигалактуроназы имеют оптимум действия при рН от 3,4 до 6,0 и температуре 30-50°С. Некоторые грибные экзо-ПГ активируются кобальтом, ингибируются щелочноземельными, тяжелыми металлами и ЭДТА.
Экзополигалактуроназы входят в состав пектолитических и цитолитических ферментных препаратов из культур микроорганизмов (Пектаваморина, Пектофоетидина, Целлюлазы-100, Поликанесцина, Ультразима, Винфлоу и др.).
Для промышленного производства ферментных препаратов пектолитических ферментов, которые являются комплексными, в качестве продуцентов используют в основном микроскопические (плесневые) грибы, в частности, грибы рода Aspergillus: A. niger, A. wentii, A. oryzae. Бактериальные ферменты в промышленных масштабах не производятся.
Растительные полигалактуроназы, по-видимому, похожи на грибные полигалактуроназы. Они обнаружены в широком спектре плодов и овощей: помидорах, авокадо, редисе, огурцах, яблоках, грушах, цитрусовых и др. Все они проявляют активность при естественных рН плодов.
Применение препаратов пектолитических ферментов в промышленности достаточно обширно. Они используются при производстве фруктовых соковых концентратов и экстрактов, при осветлении соков и вин, при производстве фруктовых и овощных пюре и нектаров.
Целлюлолитические ферменты
Целлюлоза является одним из наиболее трудно гидролизуемых природных полимеров. В организме высших животных и человека не синтезируются ферменты, гидролизующие целлюлозу. Биодеградацию целлюлозы осуществляют ферменты микроорганизмов. Микрофлора толстого кишечника человека ферментирует целлюлозу овощей и фруктов полностью. Более грубая целлюлоза, например, входящая в препараты пищевых волокон, расщепляется на 0–70%.
В гидролизе целлюлозы участвуют три основных вида ферментов. Эндо-β-1,4-глюканазы (КФ3.2.1.4) катализируют неупорядоченное расщепление целлюлозных молекул на крупные фрагменты. При действии экзо-β-1,4-глюканазы, или целлюлобиогидролазы (К.Ф.3.2.1.91) от нередуцирующего конца целлюлозных молекул или их ферментов отщепляется целлобиоза. Целлобиозы, или β-глюкозидазы (К.Ф.3.2.1.21) катализируют гидролиз целлобиозы и, с меньшей скоростью, небольших целлоолигосахаридов, с образованием глюкозы. Некоторые микроорганизмы синтезирую экзо-β-1,4-глюкозидазу (КФ3.2.1.74), под действием которой от нередуцирующего конца целлюлозных субстратов отщепляется глюкоза.
Индивидуальные эндо- и экзоглюканазы способны расщеплять нативную целлюлозу, однако в природе этот процесс происходит обычно под действием комплекса ферментов.
Целлюлазные комплексы микроорганизмов и высших базидиальных грибов включают до 20 ферментных белков, среди которых, как правило есть и эндо-, и экзо-ферменты.
Гидролиз целлюлозы ассоциированными бактериальными целлюлазами имеет место в рубце жвачных животных. В рубцовой жидкости лишь около 5% целлюлаз находится в свободном состоянии, остальная часть представлена ассоциатами. В гидролизе целлюлозы участвуют различные бактерии, населяющие рубец. За 6-8 ч пребывания в этом отделе желудка целлюлоза расщепляется на 40-50%.
Полнота гидролиза целлюлозы зависит от ряда факторов, в числе которых следующие: степень кристалличности субстрата, величина его удельной поверхности, состав ферментативного комплекса, используемого для гидролиза, и свойства его компонентов.
Нативная целлюлоза имеет очень прочную структуру и трудно гидролизуется. При исследовании гидролиза образцов целлюлозы различной степени кристалличности найдена обратная зависимость скорости гидролиза от процента кристалличности. Для увеличения доступности целлюлозы действию ферментов ее подвергают измельчению. При этом снижается размер частиц, увеличивается удельная поверхность субстрата и доля аморфной части. При сильном механическом воздействии может быть даже снижена степень полимеризации целлюлозы. Скорость гидролиза целлюлозы прямо пропорциональна величине удельной поверхности, она увеличивается по мере снижения размера частиц и степени полимеризации целлюлозы.
Микроорганизмы синтезируют целлюлазные комплексы, различающиеся по способности гидролизовать целлюлозу с высокой степенью кристалличности. Так называемые «неполноценные» комплексы хорошо гидролизуют аморфную целлюлозу, а в кристаллической целлюлозе расщепляют лишь ее аморфную фракцию (2-5%). Резкое снижение активности «неполноценных» комплексов по отношению к «полноценным» наблюдается при возрастании степени кристалличности субстрата до 60-70%.
Полноценные целлюлазные комплексы обязательно содержат эндоглюканазы, способные прочно сорбироваться на субстрате.
Чем выше коэффициент распределения, тем выше реальная концентрация фермента на поверхности субстрата и скорость гидролиза. Наблюдается прямая пропорциональная зависимость скорости гидролиза кристаллического субстрата от количества эндоглюканазы, сорбированной на его поверхности
В полноценном целлюлазном комплексе, где, как правило, присутствует эндо-фермент с высокой сорбционной способностью, целлобиогидролаза может и не обладать этим свойством. При этом наблюдается синергизм действия целлобиогидролазы и эндоглюканазы: активность комплекса ферментов выше суммы активностей его составляющих. Синергизм объясняется тем, что эндо фермент путем механохимического воздействия подготавливает субстрат для экзо-фермента, а также последовательностью реакций расщепления целлюлозы, катализируемых эндо- и экзоглюканазами, по схеме:
| |||
| |||

 Целлюлоза Целлодекстрины
Целлюлоза Целлодекстрины
|

 Целлобиоза Глюкоза
Целлобиоза Глюкоза
Синергизм действия может наблюдаться в различных комбинациях эндо- и экзо-ферментов (эндо-эндо, эндо-экзо, экзо-экзо), но в любом случае одна из целлюлаз значительно отличается от другой по способности адсорбироваться на субстрате. Ферменты, близкие по сорбционной способности, при соединении не проявляют синергизма. Синергический эффект целлюлаз значителен: степень расщепления субстрата увеличивается в 2,5-2,8 раза.
Для гидролиза целлюлозы используются комплексные ферментные препараты, выделяемые из культур микроскопических грибов и актиномицетов и обладающие эндоглюканазной, целлобиогидролазной и целлобиазной активностью. Отдельные компоненты целлюлазных комплексов грибов и актиномицетов проявляют наибольшую активность при рН от 3,7 до 5,5, а комплексы в целом – при рН 4,5-5,5. Оптимальная температура действия отдельных компонентов – от 45 до 80°С, комплексов – 50-60° С. Некоторые высшие базидиомицеты синтезируют целлюлазы с оптимумом при рН З.
Многие целлюлазы являются углеводсодержащими белками, углеводная часть может составлять до 90% молекулярной массы. Углеводная часть выполняет якорную функцию, способствуя сорбции фермента на субстрате. Сорбция по сродству необходима, поскольку в рН-зоне активности целлюлазы имеет незначительный заряд (рН-оптимумы близки к ИЭТ). Возможно, углеводная часть обеспечивает скольжение фермента в фибриллярных структурах целлюлозы. Это существенно, поскольку целлюлазы осуществляют сотни и тысячи каталитических актов, не покидая поверхности одной целлюлозной молекулы.
Углеводная часть целлюлаз защищает белок от действия денатурирующих агентов и от протеолиза.
Конверсия целлюлозы в природных биоценозах сопряжена с деструкцией гемицеллюлозы и лигнина. При культивировании грибов на древесных субстратах в первую очередь разлагается гемицеллюлоза, после удаления ксилана увеличивается скорость гидролиза целлюлозы. Ксилазы и целлюлазы проявляют синергизм, что объясняется последовательностью их действия при гидролизе смешанного субстрата, где целлюлоза экранирована гемицеллюлозой.
Применение целлюлолитических ферментов представляет большой интерес, т.к. ферментативный гидролиз целлюлозосодержащих материалов (древесина, торф, сельскохозяйственные и городские отходы) может обеспечить получение различных биотехнологических продуктов (глюкозы, этанола, ацетона, микробной биомассы).








